Browse Articles
Conditioning Medicine
International bi-monthly journal of cell signaling, tissue protection, and translational research.
The microRNA miR-21 conditions the brain to protect against ischemic and traumatic injuries
Time:2017-12-11
Number:12034
Author Affiliations

Conditioning Medicine, 2017. 1(1):35-46.
Abstract
Ischemic and traumatic injuries to CNS remain leading causes of death and disability worldwide, despite decades of research into risk factors, therapies, and preventative measures. Recent studies showed that CNS injuries significantly alter the cerebral microRNAome that impact the secondary brain damage as well as plasticity and recovery. Many microRNA based therapies are currently in various clinical trials for different pathologic conditions indicating their therapeutic potential. In the present review, we discuss the role of miR-21 in acute CNS injuries which is currently thought to be a potent neuroprotective microRNA. We emphasize on the potential of miR-21 in promoting cell and tissue survival and preventing inflammation and apoptosis. We also discussed the role of miR-21 in conditioning the brain to promote ischemic tolerance. Finally, we discussed some of the challenges and difficulties to develop miR-21 as a neuroprotective therapy in humans.
Keywords: Stroke; CNS injury, therapy; miR-21; neuroprotection
Abstract
Ischemic and traumatic injuries to CNS remain leading causes of death and disability worldwide, despite decades of research into risk factors, therapies, and preventative measures. Recent studies showed that CNS injuries significantly alter the cerebral microRNAome that impact the secondary brain damage as well as plasticity and recovery. Many microRNA based therapies are currently in various clinical trials for different pathologic conditions indicating their therapeutic potential. In the present review, we discuss the role of miR-21 in acute CNS injuries which is currently thought to be a potent neuroprotective microRNA. We emphasize on the potential of miR-21 in promoting cell and tissue survival and preventing inflammation and apoptosis. We also discussed the role of miR-21 in conditioning the brain to promote ischemic tolerance. Finally, we discussed some of the challenges and difficulties to develop miR-21 as a neuroprotective therapy in humans.
Keywords: Stroke; CNS injury, therapy; miR-21; neuroprotection
Introduction
Various classes of non-coding RNAs (ncRNAs) that represent >98% of the transcriptional output in humans are considered as major controllers of transcription and translation (Cech and Steitz, 2014). The ncRNAs are diverse in size. They can be small like microRNAs (miRNAs) and piwi-interacting RNAs or medium-sized like small nucleolar RNAs and small Cajal body-specific RNAs or big like long noncoding RNAs (lncRNAs) and telomere-associated RNAs (Vemuganti, 2013). The ncRNAs are also very diverse in their function (Chandran et al., 2017). Ribosomal RNAs and transfer RNAs are the most studied ncRNAs that control translation. Whereas, miRNAs which are ~22 nucleotides long also control translation by binding to 6-8 nucleotide complementary seed sequences in the 3’ untranslated regions (3’-UTRs) of mRNAs (Lee et al., 2002; Diederichs and Haber, 2007). The lncRNAs are known to modulate transcription by binding to other RNAs, chromatin modifying proteins, transcription factors and DNA (Kung et al., 2013).
Of the various classes of ncRNAs, miRNAs are the most studied. New miRNAs are continuously being discovered in all species and as of now 2,588 mature miRNAs were discovered in humans and 1,915 in mouse (http://www.mirbase.org/). The miRNAs are transcribed from specific genes by RNA polymerase II as several hundred base pairs of stem-loop structures called primary miRNAs (pri-miRNAs) (Lee et al., 2002). A pri-miRNA will be cleaved in the nucleus by RNase Drosha in association with a protein called DiGeorge Syndrome Critical Region 8 (DGCR8) to release 1 to 6 precursor miRNAs (pre-miRNAs) (Lee et al., 2003). The pre-miRNAs are ~80 nucleotide long hairpin structures that will be transported to cytosol by exportin-5 and cleaved by the RNase III Dicer to release mature miRNAs which are 18-24 nucleotides long (Diederichs and Haber, 2007).
Recent studies have shown that miRNAs play crucial roles in vertebrate development and disease progression including heart failure, diabetes, cancer, and various brain and kidney pathologies (Kantharidis et al., 2011; Jansson and Lund, 2012; Jimenez-Mateos and Henshall, 2013; Olson, 2014). Several miRNA-based therapies also transitioned from preclinical studies to clinical trials. These include antimiR-122 for hepatitis C, antimiR-155 for cutaneous T-cell lymphoma, miR-34 mimic for solid tumors, miR-16 mimic for mesothelioma, miR-29 mimic for scleroderma (Rupaimoole and Slack, 2017). Preclinical studies showed that miR-122 is required for the replication of hepatitis C virus in liver (Baek et al., 2014). Following this lead, miravirsen (miR-122 antagonist) is currently in a phase 2a clinical trial and the initial results showed decreased viral load of hepatitis C for up to 14 weeks after the last treatment (Janssen et al., 2013; van der Ree et al., 2014). MiR-34a is a tumor suppressor miRNA that is often downregulated or lost in solid tumors, including breast, colorectal, liver, lung, and prostate cancer (Saito et al., 2015; Adams et al., 2016). Transfection with a miR-34a mimic decreased invasion/migration in cultured human hepatocellular carcinoma cells (Li et al., 2009) and promoted apoptosis in multiple myeloma cell lines (Di Martino et al., 2012). Furthermore, mouse models of prostate cancer and diffuse B-cell lymphoma showed tumor regression and improved survivorship after intravenous treatment with miR-34a mimic (Liu et al., 2011; Craig et al., 2012). Encouragingly, MRX34 (miR-34a mimic encapsulated in liposomes) recently met acceptable safety standards and showed potential antitumor activity in a phase I clinical trial (Beg et al., 2017). These clinical trials show the potential of miRNA-based therapies for cancers as well as other conditions like stroke with not many available drugs (Mozaffarian et al., 2016).
miRNAs modulate secondary neuronal damage following acute injuries to CNS
Traumatic and ischemic insults to the brain or spinal cord lead to significant neurological deficits that are exacerbated by secondary neuronal damage mediated by many synergistic pathophysiologic mechanisms that include excitotoxicity, inflammation, oxidative stress and apoptosis (Dreier, 2011; Dirnagl, 2012; Li et al., 2017). Recent studies showed that both ischemic and traumatic injuries to CNS are associated with altered cerebral miRNA expression profiles (Vijayan and Reddy, 2016; Di Pietro et al., 2017). Many preclinical studies also showed that secondary brain damage and neurological dysfunction can be curtailed and/or restorative mechanisms like neurogenesis and angiogenesis can be induced by manipulating specific miRNAs (Lou et al., 2012; Liu et al., 2013b; Zeng et al., 2014). In this section, we discuss the role of miRNAs after CNS injury, using stroke as a specific example. Role of miRNAs after traumatic brain injury (TBI) and spinal cord injury (SCI) is discussed at a later stage in the manuscript.
Cerebral miRNA expression profiles were shown to be significantly altered as early as 30 minutes and as late as 3 days of reperfusion following focal or global cerebral ischemia in rodents (Jeyaseelan et al., 2008; Dharap et al., 2009; Yuan et al., 2010). Pathway analysis showed that ischemia leads to upregulation of miRNAs that target pro-survival mRNAs and down-regulation of miRNAs that target pro-apoptotic and/or pro-inflammatory mRNAs (Jeyaseelan et al., 2008; Liu et al., 2010a; Hunsberger et al., 2012). This leads to an unfavorable environment for cellular survival in the ischemic brain.
Many preclinical studies evaluated the significance of modulating specific miRNAs to decrease secondary brain damage and/or to induce plasticity/regeneration after stroke. The miR-145 was reported to be significantly upregulated in rat brain following transient focal ischemia, and treatment with antagomiR-145 was shown to decrease infarction (Dharap et al., 2009). This neuroprotective effect was observed to be mediated by de-repression of miR-145 target superoxide dismutase-2 (SOD2), which is an antioxidant protein that alleviates oxidative stress in the ischemic brain (Dharap et al., 2009). Many other miRNAs were also shown to modulate post-ischemic pathology. Notable examples are let-7f, miR-23a and miR-497. Improved neurological outcome and decreased infarct volume after focal ischemia was shown by inhibiting let-7f (targets insulin-like growth factor 1), miR-23a (targets X-linked inhibitor of apoptosis) and miR-497 (targets pro-apoptotic Bcl2) (Yin et al., 2010; Siegel et al., 2011; Selvamani et al., 2012).
Due to the redundancy of seed sequences and presence of binding sites for multiple miRNAs in the 3’UTRs, miRNAs act in concert to affect specific pathways. For example, several miRNAs modulate post-ischemic inflammation by targeting nuclear factor kappa B (NF-κB) pathway (Buchan et al., 2000; Xu et al., 2012). When rats were transfected with an adenoviral vector overexpressing miR-22, its target nuclear receptor coactivator 1 (NCOA1; a NF-κB coactivator) was repressed leading to neuroprotection after stroke (Yu et al., 2015). MyD88, a protein adaptor for receptors that mediate nuclear translocation of NF-κB is targeted by miR-203 (Yang et al., 2015). Treatment with a miR-203 mimic was shown to silence NF-κB signaling resulting in decreased infarct volume, less edema and improved motor function recovery after stroke in mice (Yang et al., 2015). AntagomiR-181a administered either intravenous or intracerebroventricular was protective against ischemic injury in mice, likely due to degradation of GRP78, which is a NF-κB coactivator (Ouyang et al., 2012; Xu et al., 2015).
miRNAs and ischemic tolerance in the brain
Hibernating animals survive without brain damage for months with very low oxygen and glucose and this physiologic adaptation makes them resistant to stroke-induced brain damage (Zhou et al., 2001; Dave et al., 2006). Several studies have implicated miRNAs in the torpor response in different organs including liver, heart and skeletal muscle of hibernating animals including ground squirrels, bats and Dromiciops(Yuan et al., 2015; Hadj-Moussa et al., 2016; Wu et al., 2016). The brain of torpid hibernating brown bats showed altered miRNA expression when compared to non-torpid littermates (Biggar and Storey, 2014). Notably, they showed altered expression of 10 miRNAs (2 decreased and 8 increased) which target the translation of proteins that modulate focal adhesion and axon guidance (Biggar and Storey, 2014). We will discuss the therapeutic potential of miR-21 which is one of the upregulated miRNAs observed in this study. Ischemic tolerance can be induced in many species by a short duration ischemic insult which prepares the organs for a subsequent long duration ischemic event (Liu et al., 1992; Stetler et al., 2014; Varga et al., 2014). This preconditioning (PC) effect was shown to be associated with significantly altered miRNA expression profiles in rats, mice and gerbils. Following ischemic PC induced by a 10 min middle cerebral artery occlusion (MCAO) in adult rats, several miRNAs that target neuroprotective pathways were shown to be downregulated while those that target cell death pathways were upregulated in cerebral cortex from 6h to 3 days (Dharap and Vemuganti, 2010). Interestingly, miR-21 was the miRNA that showed a consistent induction (from 6h to 3 days) and also the highest fold increase in rat brain after ischemic PC (Dharap and Vemuganti, 2010). A study of in vitro ischemic PC using oxygen-glucose deprivation (OGD) in rat hippocampal neurons also showed miR-21 upregulation at 1 day of re-oxygenation (Keasey et al., 2016). Altered miRNA expression profiles were also shown in the mouse brain from 3h to 3 days following ischemic PC induced by a short duration MCAO (Keasey et al., 2016; Lusardi et al., 2010). Suppression of miR-132 and subsequent induction of the transcriptional repressor methyl CpG binding protein 2 (MeCP2) were implicated as the major mediator of PC in mouse brain (Lusardi et al., 2010). These authors showed that MeCP2 knockout mice will not develop ischemic tolerance when subjected to PC.PC induced by a short duration global ischemia in gerbils altered the expression of several miRNAs for up to 6 months, although the consequence of this was not evaluated further (Sun et al., 2015). Whereas, Lee et al. (2010) attributed the PC-induced ischemic tolerance to upregulation of miR-200 leading to degradation of its target prolyl hydroxylase 2, which is a part of the pathway that modulates hypoxia-inducible factor 1α (Ratcliffe et al., 2017).
miR-21 as a therapeutic candidate for CNS injury
The previous sections indicated that stroke alters many miRNAs and several of them can serve as biomarkers to identify a disease state as well as serve as therapeutic targets to protect the brain. Of these, miR-21, one of the first miRNAs isolated from mammalian tissue, stands out as a strong candidate for translation to human stroke therapy (Lagos-Quintana et al., 2002; Krichevsky and Gabriely, 2009; Kumarswamy et al., 2011). Putative functions of miR-21 have been investigated in many pathologies including cardiac ischemia, hepatic fibrosis and different types of cancers (Pan et al., 2010; Zhang et al., 2013b; Gu et al., 2015). Additionally, several predicted gene targets of miR-21 have been validated (Cheng et al., 2010; Buscaglia and Li, 2011). Importantly, miR-21 represses the translation of proteins that promote apoptosis and inflammation and hence helps the post-ischemic outcome (Xu et al., 2014).
Anti-apoptotic and anti-inflammatory functions of miR-21
The best studied targets of miR-21 are programmed cell death 4 (PDCD4), Fas ligand (FasL) and phosphatase and tensin homologue (PTEN) (Fig. 1). All of these are strongly implicated in promoting apoptosis and inflammation (Lu et al., 2008; Sayed et al., 2010; Sheedy et al., 2010; Das et al., 2014; Choi et al., 2015; Wu et al., 2015; Yang et al., 2016a). The dual role of these proteins emphasizes that apoptosis and inflammation potentiate each other in a feedback loop. PDCD4 promotes apoptosis by binding to the translational initiation factor eIF4a and arresting translation of activator protein 1 (AP-1) (Yang et al., 2001; Loh et al., 2009). FasL is expressed on the surface of immune cells and binds to Fas death receptor to promote apoptosis via caspase cleavage and/or p53 activation (Benchimol, 2001; Broughton et al., 2009). PTEN dephosphorylates phosphatidylinositol 1,4,5-trisphosphate (PIP3) to form phosphatidylinositol 4,5-bisphosphate (PIP2), preventing PIP3-mediated activation of the kinase Akt, resulting in caspase cleavage (Tu et al., 2013; Yang et al., 2014). Loss of PDCD4, FasL and PTEN contributes to increased survival of cancer cells (Chen et al., 2003; Viard-Leveugle et al., 2003; Peng et al., 2016). Many studies implicated miR-21 as an oncomiR since its enrichment in primary tumors negatively correlates with the expression of these proteins (Chen et al., 2008; Li et al., 2013a; Shang et al., 2015). However, it is key to bear in mind that this anti-apoptotic function which is deleterious for cancer is beneficial in conditions of acute brain injury by promoting cell survival.
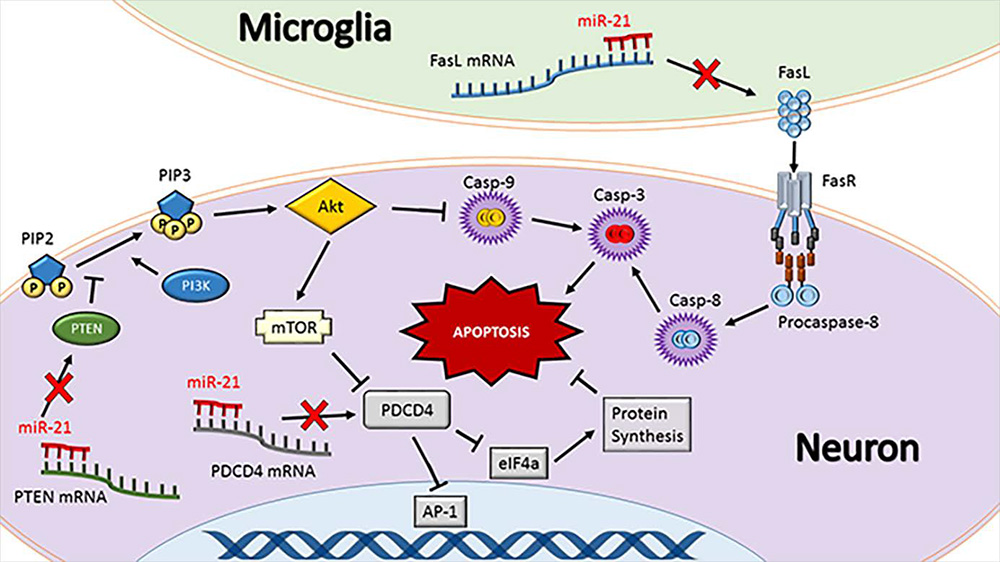
In a new window | Download PPT
Figure 1: miR-21 regulates apoptosis. Fas ligand (FasL), programmed cell death protein 4 (PDCD4), and phosphatase and tensin homolog (PTEN) are the best-characterized targets of miR-21. These proteins synergistically promote apoptosis. FasL is either membrane-bound or soluble and physically binds to the Fas death receptor (FasR) to initiate a caspase cleavage cascade resulting in apoptosis. PTEN is a tumor suppressor and a negative regulator of PI3K that phosphorylates PIP2 to form PIP3 that activates Akt pathway leading to cell death. As such, prevention of PTEN by miR-21 can lead to better cell survival following conditions like stroke and TBI. PI3K and Akt also modulate caspase-9 cleavage and hence, PTEN inhibition leads to prevention of apoptosis. Furthermore, PI3K/Akt activates mammalian target of rapamycin (mTOR) that inhibits PDCD4 activity. PDCD4 causes a protein expression bottleneck by inhibiting eukaryotic initiation factor 4a (eIF4a) and the transcription factor activator protein 1 (AP-1). Preventing eIF4a activity causes endoplasmic reticulum stress and inhibiting AP-1 prevents expression of beneficial genes such as vascular endothelial growth factor (VEGF). As inhibition of FasL, PTEN or PDCD4 together is a powerful strategy to prevent apoptosis and hence miR-21 is an attractive therapeutic to protect the post-injury brain. Tipped arrows indicate activation and blunted arrows indicate inhibition.
The anti-inflammatory effect of miR-21 has been demonstrated in models of acute inflammation, peritonitis, cardiovascular disease and kidney injury (Feng et al., 2014; Toldo et al., 2014; Jia et al., 2015; Barnett et al., 2016; Zhang et al., 2016). The miR-21 target PDCD4 in particular has been shown to activate NF-κB transcription leading to inhibition of the anti-inflammatory cytokine IL-10 ( Yang et al., 2001; Young et al., 2010). PTEN/Akt signaling is required for site-directed immune cell migration due to control of its downstream factors mammalian target of rapamycin (mTOR) and FoxO1, which modulate cell growth and metabolism (Li et al., 2000; Hedrick et al., 2012; Zhang et al., 2013a; Xie et al., 2014). Additionally, PTEN knockout mice are resistant to pneumonia infection (Schabbauer et al., 2010). The membrane bound isoform of FasL promotes neutrophil recruitment acting as a chemoattractant for these cells (Hohlbaum et al., 2000; Dupont and Warrens, 2007).
In addition to FasL, PDCD4, and PTEN, miR-21 targets many mRNAs that encode pathologically relevant pro-inflammatory proteins. The transcription factor signal transducer and activator of transcription 3 (STAT3) is a validated target of miR-21 (Wang et al., 2015). Once activated by transmembrane receptor kinases, STAT-3 translocate into the nucleus and transcribes many genes including SOD2, Bcl family and intracellular adhesion molecule 1 (Dauer et al., 2005; Yu et al., 2009). All these are known to modulate oxidative stress, inflammation and apoptosis. Interestingly, colon and skin cancer cells implicate miR-21/PTEN/Akt axis and STAT3 in a feedback regulatory loop such that STAT3 knockdown prevents miR-21 expression, and overexpression of miR-21 increases IL-6 levels which in turn activates STAT3 (Iliopoulos et al., 2010; Lu et al., 2015).
Clinical studies implicated that miR-21 silencing could be therapeutically beneficial in conditions like chronic inflammatory diseases, diabetic nephropathy and renal fibrosis (Zhang et al., 2011; Wang et al., 2014; McClelland et al., 2015). Increased levels of miR-21 in the blood of patients with chronic kidney fibrosis suggest that miR-21 might be a robust biomarker of this disease (Glowacki et al., 2013). MiR-21 expression was also shown to be elevated significantly in blood samples from patients suffering with chronic cardiopulmonary diseases, cardiac fibrosis, idiopathic pulmonary fibrosis and coronary heart disease (Liu et al., 2010b; Li et al., 2015; Lorenzen et al., 2015). A seemingly contradictory finding attributed to miR-21 is silencing its target SMAD7, a signaling molecule that binds to the intracellular region of the transforming growth factor beta receptor (TGFβR), and thus prevents phosphorylation of SMAD2 and SMAD3 (Wang et al., 2014; Choi et al., 2016). When SMAD2/3 are phosphorylated, they translocate into nucleus to promote transcription of extracellular matrix proteins such as collagen I and fibronectin (Meng et al., 2015b). The role of miR-21 in controlling SMAD7 was thought to be important in various conditions like hepatic fibrosis, atrial fibrosis, pulmonary fibrosis and carcinoma-associated fibroblast formation (Li et al., 2013b; He et al., 2016; Kwon et al., 2016; Yang et al., 2016b).
Role of miR-21 in stem cell integrity and angiogenesis
Embryonic and new born mouse brains were shown to be enriched with miR-21 that correlates with SOX2 levels till postnatal day 7 (Polajeva et al., 2012). Furthermore, miR-21 and SOX2 levels showed similar pattern of induction in mouse brain tumor samples, and treatment with antagomiR-21 significantly reduced SOX2 levels in both mouse and human glioma cells (Polajeva et al., 2012). As SOX2 is a transcription factor required for neural stem cell pluripotency, this study indicates the role of miR-21 in stem cell pluripotency, brain development and glioma expansion. A recent study showed increased proliferation of neural progenitor cells cultured in hypoxic conditions that can be attributed to activation of the miR-21/PTEN/Akt axis (Chen et al., 2017).
Increased levels of miR-21 are associated with better integrity of cardiac stem cells (CSC) and their proliferation via its target PTEN. Rat CSCs transfected with miR-21 mimic were robustly protected against lethal H2O2 exposure which was associated with decreased PTEN levels resulting in increased Bcl:Bax ratio, decreased cleaved caspase-3 levels and fewer apoptotic cells (Hori and Nishida, 2009; Deng et al., 2016). CSCs from both mice and rats showed increased proliferation and trans-well migration after transfection with miR-21 mimic (Zhou et al., 2016; Shi et al., 2017). Improved proliferation/migration of stem cells was mimicked and attenuated respectively, by small-molecule inhibitors of PTEN and PI3K, suggesting that PTEN degradation is a key mechanism of miR-21-mediated proliferation/migration of CSCs (Shi et al., 2017).
Exosomes from stem cell are known to contain high levels of pro-survival proteins, RNAs and intact mitochondria (Lai et al., 2011; Phinney et al., 2015). Exosomes derived from human endometrial mesenchymal stem cells and cardiomyocyte progenitor cells were found to be enriched with miR-21. Rat cardiomyocytes cultured in exosome conditioned medium were resistant to oxidative stress-mediated apoptosis in vitro and this protection was attributed to uptake of miR-21 and degradation of its target PDCD4 (Xiao et al., 2016). A subsequent study showed that exosome-conditioned rat cardiomyocytes exhibited improved engraftment and survival after transplantation into the ischemic rat heart, and this improvement was attributed to prevention of PTEN translation by miR-21 (Wang et al., 2017). Together, these studies support a pro-recovery role of miR-21 after an injury to promote plasticity after stroke (Barkho and Zhao, 2011; Goritz and Frisen, 2012).
Role of miR-21 in stroke
Preclinical models show a highly phasic pattern of miR-21 expression after ischemic insults. Rat cortical neurons subjected to OGD showed induction of miR-21 by 1 day (Ziu et al., 2011). However, in microglia subjected to hypoxia, miR-21 was downregulated leading to induction of its target Fas ligand that was thought to promote apoptosis (Zhang et al., 2012). Adult rats subjected to global ischemia or embolic focal ischemia also showed induction of miR-21 at 1 day of reperfusion (Deng et al., 2013; Liu et al., 2013a; Li et al., 2016b). In the rat MCAO model of focal ischemia, miR-21 upregulated significantly in the penumbra from 2 to 7 days post reperfusion (Buller et al., 2010). These studies indicate that miR-21 might be a player in promoting neuroprotection as well as plasticity and recovery after stroke.
MiR-21 was shown to inhibit apoptosis by preventing FasL protein expression after OGD (Buller et al., 2010). In cultured rat cortical neurons subjected to OGD, FasL levels and apoptosis were curtailed by transfection with miR-21 mimic and induced by transfection with antagomiR-21 (Buller et al., 2010). A subsequent study showed that when rat hippocampal neurons were cultured in conditioned medium from rat microglia subject to OGD, there was an induction of FasL and increased apoptosis (Zhang et al., 2012). Furthermore, transfecting the microglia with a miR-21 mimic before the exposure to OGD attenuated FasL expression and prevented neuronal death in hippocampal neurons cultured with the conditioned medium (Zhang et al., 2012). Peripheral blood samples from ischemic and hemorrhagic stroke patients collected at an acute stage (12 to 24h after stroke) showed significantly lower miR-21 levels compared to healthy age-matched controls (Zhou and Zhang, 2014; Wang et al., 2016). Interestingly, elevated miR-21 in the peripheral blood correlated with increased incidence of stroke-associated infections and decreased serum interferon gamma (IFNγ) levels (Lin et al., 2016). This is attributes a putative anti-inflammatory effect to miR-21.
Role of miR-21 in traumatic brain injury
Neuronal death and neurological dysfunction after TBI is also synergistically mediated by apoptosis, inflammation and oxidative stress similar to stroke (Masel and DeWitt, 2010). However, there are certain key differences between TBI and stroke pathologies. For example, post-TBI lesions exhibit significant diffuse axonal injury and increased necrosis unlike post-stroke infarcts (Vieira et al., 2016). Furthermore, TBI is highly associated with hemorrhages that contribute to the secondary brain damage (Nolan, 2005). Despite these differences, preclinical studies show similar patterns of miR-21 expression in TBI and stroke. In rats, fluid percussion injury (FPI) upregulated miR-21 expression in the contused cortex at 1 to 7 days after the injury (Lei et al., 2009; Ge et al., 2014; Ge et al., 2015). Whereas, controlled cortical impact (CCI) injury in rats was shown to down-regulate miR-21 levels at 3h with subsequent upregulation from 1 to 3 days after the injury (Redell et al., 2011). Mice subjected to CCI injury also showed upregulation of miR-21 from 6h to 7 days post injury (Sandhir et al., 2014; Meissner et al., 2016). The miR-21 was reported to be enriched in the extracellular vesicles isolated from the contused hemisphere of mice at 7 days after CCI injury (Harrison et al., 2016). Expression patterns of miR-21 are different in aged and young mice, such that miR-21 levels returned to sham levels by 1 day after injury in aged mice, while they stayed elevated till 7 days post injury in young mice (Sandhir et al., 2014). Compared to young mice, aged mice showed a higher degree of secondary brain damage and increased expression of the miR-21 target proteins PTEN, PDCD4 and tissue inhibitor of metalloproteinase 3 (TIMP3) (Sandhir et al., 2014). Intracerebroventricular injection of miR-21 mimic to rats after FPI was shown to suppress PTEN expression with corresponding increased levels of p-Akt, decreased TUNEL staining, lower Bcl2/Bax ratio and decreased cleaved caspase-3 levels that led to reduced lesion size and curtailed edema and improved behavioral outcome (Ge et al., 2014; Ge et al., 2015). Thus, miR-21 was shown to inhibit apoptosis by promoting the degradation of PTEN. The miR-21-PTEN-Akt axis was also explored in cultured rat cortical neurons subjected to scratch injury (Morrison et al., 2011). Neurons transfected with miR-21 mimic 1 day before scratch injury showed PTEN degradation, increased Bcl2/Bax ratio, decreased TUNEL staining and decreased levels of cleaved caspase-3 and 9 similar to that observed in the in vivo TBI models (Han et al., 2014). Apoptosis after TBI may be modulated by PDCD4, which inversely correlated with mir-21 expression in CCI-contused rats and mice (Redell et al., 2011; Sandhir et al., 2014). All these studies indicate that induction of miR-21 is a neuroprotective adaptation to protect the brain after TBI.
After TBI, miR-21 seems to be playing a neurorestorative role as well. At 3 days post-FPI, rats treated with miR-21 mimic showed induction of claudin-5 and occludin in the injury lesion and boundary (Ge et al., 2015). At 7 days post-FPI, miR-21 treated rats showed increased microvascular density and vascular endothelial growth factor (VEGF) expression (Ge et al., 2014). At both 3 and 7 days post-FPI, edema was decreased, and expression of Ang1 and Tie2 was increased compared to controls; however the mechanism of miR-21-mediated effects on Ang1/Tie2 axis was not evaluated in detail (Ge et al., 2014; Ge et al., 2015). These studies in general indicate that miR-21 promotes blood-brain barrier (BBB) integrity and angiogenesis after TBI. Many targets of miR-21 are pro-inflammatory and pro-apoptotic. When rat brain microvascular endothelial cells transfected with miR-21 mimic were subjected to scratch injury, expression of the pro-inflammatory TNFα, IL-6 and NF-κB decreased and the expression of the anti-inflammatory IL-10 increased (Ge et al., 2016). Furthermore, the scratch injured cells transfected with miR-21 mimic showed decreased apoptosis (as measured by Annexin V and cleaved caspase-9 staining) and elevated levels of claudin-5, occludin, Ang1, and Tie2 that might promote endothelial cell function (Ge et al., 2016).
Role of miR-21 in spinal cord injury
SCI causes extensive sensory and motor deficits that are synergistically mediated by several secondary injury mechanisms that include excitotoxicity, inflammation, oxidative stress and apoptosis (Silva et al., 2014). Despite these mechanistic similarities, SCI pathology is distinct from stroke pathology and poses its own set of challenges (Oyinbo, 2011). The average SCI patient is younger than the average stroke patient and hence the post-injury period of survival is longer. SCI has an extended chronic phase that is characterized by the formation of a perilesional glial scar composed of astrocytes and extracellular matrix that prevents axonal growth (Fawcett and Asher, 1999).
Rats subjected to SCI showed increased miR-21 expression between 3 and 35 days post-injury (Liu et al., 2009; Bhalala et al., 2012; Yunta et al., 2012; Hui et al., 2013). A single bolus of intravenous administration of the omega-3 fatty acid docosahexaenoic acid (DHA) that improves learning, memory, and motor function recovery after SCI (Lim et al., 2013) increased miR-21 levels leading to decreased levels of its target PTEN (Liu et al., 2015). Furthermore, neuronal cultures treated with DHA showed increased neurite growth which was attenuated by antagomiR-21 treatment (Liu et al., 2015). Intrathecal administration of antagomiR-21 after SCI led to increased expression of FasL and PTEN, bigger lesion size and less recovery of the hind limb function (Hui et al., 2013). A recent study showed that in rats subjected to weight drop SCI, treatment with antagomiR-21 increased TIMP3 expression and decreased vascular density, whereas treatment with either miR-21 mimic or TIMP3 siRNA induced MMP2 and MMP9 expression and improved capillary network formation (Hu et al., 2016).
Though the mechanism is unclear, miR-21 was shown to play a role in ameliorating astroglial hypertrophy. Lentivirus-mediated overexpression of miR-21 decreased GFAP expression and cell size in primary mouse astrocytes (Sahni et al., 2010). Transgenic mice that overexpress miR-21 showed decreased GFAP density and thinner astrocytic processes at 14 and 35 days after SCI compared to wild-type controls (Bhalala et al., 2012). This group further demonstrated that transgenic mice that express a miR-21 sponge show persistent astrocytic activation and hypertrophy up to 35 days post-SCI. Mice expressing the miR-21 sponge showed increased axon density across the SCI lesion, which is contradictory to the evidence presented above that miR-21 antagonism is associated with poor outcome after SCI (Hui et al., 2013). The authors justified this discrepancy by hypothesizing that the activated, hypertrophic astrocytes in the miR-21 sponge expressing mice prevented formation of the glial scar (Bhalala et al., 2012). Overall, these studies indicate that overexpression of miR-21 at an acute stage after SCI is potentially therapeutic, but inhibition of miR-21 during the chronic phase after SCI might promote recovery.
Role of miR-21 in other CNS pathologies
MiR-21 is also implicated in several non-ischemic, non-traumatic CNS pathologies. It was shown to be upregulated in the cerebral cortex of autism patients (Mor et al., 2015), in the CSF of patients with viral CNS infection (Goswami et al., 2017) and in peripheral blood mononuclear cells from multiple sclerosis patients (Fenoglio et al., 2011). Studies showed that exposure of murine cortical neurospheres to high doses of alcohol repress miR-21 (Sathyan et al., 2007; Balaraman et al., 2012). However, the functional significance of miR-21 in all these conditions is not known.
Role of miR-21 in epilepsy, HIV/SIV and Parkinson’s disease (PD) was evaluated in detail. In the rat hippocampus, miR-21 was observed to be significantly down-regulated at 1 day, but upregulated from 2 to 30 days following lithium pilocarpine (LiP)-induced epilepsy (Hu et al., 2013; Risbud et al., 2011; Meng et al., 2015a; Roncon et al., 2015; Chak et al., 2016). In both juvenile and adult epileptic patients, miR-21 was reported to be upregulated in the hippocampus (Peng et al., 2013; Roncon et al., 2015). These findings corroborate earlier studies which show that suppression of hippocampal and forebrain neurotrophin 3 (NT3; a miR-21 target gene) is associated with development of status epilepticus in LiP-treated rats (Schmidt-Kastner and Olson, 1995; Mudo et al., 1996). In the brains of SIV-infected rhesus monkeys and HIV-infected humans, miR-21 was reported to be upregulated in neurons, and trafficked to microglia via extracellular vesicles (Yelamanchili et al., 2015). This neuronal upregulation of miR-21 inversely correlated with levels of its target monocyte enhancement factor 2c (MEF2c) (Yelamanchili et al., 2010) which is a transcription factor associated with potassium channel signaling required for neuronal survival and learning (Barbosa et al., 2008). In the context of HIV/SIV, miR-21-mediated decrease in MEF2c resulted in outward potassium neuronal currents, which is a sign of neuronal commitment to apoptosis (Yelamanchili et al., 2010). Cerebral mir-21 levels were observed to be markedly upregulated in the MPTP-induced PD in mice (Su et al., 2016). Mir-21 was shown to target lysosome-associated membrane protein 2 which increases the degradation of α-synuclein by promoting chaperone-mediated autophagy and thus might prevent the PD-like pathology (Xilouri et al., 2013; Su et al., 2016).
Steps to therapeutic translation
The evidence presented previously suggests that miR-21 is already an ideal candidate for translation to the clinic. However, most of these studies were conducted with either cell cultures or rodent models of stroke, TBI and SCI. For therapeutic clinical translation of miR-21, further robust and large-scale preclinical studies are needed. For example, the post-stroke therapies need to be tested following the recommendations of the Stroke Treatment Academic Industry Roundtable (STAIR) criteria that include testing both sexes, different age groups, minimal efficacious dose with no toxicity, half-life, different routes of administration, window of therapeutic opportunity, longer-term multiple functional outcomes, extensive physiological monitoring, multiple disease models and importantly randomization and blinding of the studies (Saver et al., 2009; Savitz et al., 2011). It is also important to develop a proper tissue-specific delivery strategy without the toxicity of a transfection agent and preventing the RNase-mediated degradation are important for translating a miRNA therapy to humans. Studies are underway to test encapsulating the miRNA in liposomes and attaching a targeting moiety like in dendrimers to increase tissue-specific delivery and to chemically modify a miRNA by locked nucleic acid or phosphorothioate modifications to resist the RNases (Rupaimoole and Slack, 2017). Given the pro-survival function of miR-21, the most pressing concern is to confirm that patients don’t develop tumors with miR-21 therapy (Karsy et al., 2012; Pfeffer et al., 2015; Li et al., 2016a; Li et al., 2016c). The path to therapeutic translation is clear, but needs more studies.
Summaries and Conclusions
All the above studies suggest that miR-21 is a promising candidate for therapeutic amelioration of secondary neuronal injury following acute and chronic insults to CNS. Importantly, increasing miR-21 levels might precondition the CNS to induce tolerance in case of an insult. The pro-survival effects of miR-21 in the CNS are typically attributed to inhibition of pro-apoptotic pathways controlled by the miR-21 targets PDCD4, PTEN, RECK, and FasL. Dozens of other pro-apoptotic targets of miR-21 have been verified in non-CNS paradigms (Buscaglia and Li, 2011).
Preventing apoptosis often results in decreased inflammatory burden by decreasing the amount of injured tissue requiring repair or clearance. However, miR-21 may directly affect inflammation via its gene targets PELI1, IL-12a, PTEN, FasL, and PDCD4, which regulate the NF-κB pathway. Further evidence for a directly anti-inflammatory role of miR-21 comes from investigations of peritonitis and LPS exposure, in which groups with elevated miR-21 levels survived and recovered better than the control groups (Feng et al., 2014; Barnett et al., 2016).
Encouragingly, the benefits of miR-21 are not limited to preventing damage, but are also tied to its ability to promote cell survival after injury. Neural and cardiac stem cells treated with miR-21 (via either transfection or culturing with miR-21 enriched exosomes) subjected to stressful stimuli such as hypoxia or H2O2 exposure showed decreased apoptosis and improvements in proliferation and migration, which are often considered as signs of stem cell integrity (Chen et al., 2017; Shi et al., 2017). MiR-21 was also shown to promote angiogenic recovery after SCI via its target TIMP3, which inhibits MMP2 and MMP9, and was shown to block VEGF binding to its receptor (Qi et al., 2003; Hu et al., 2016).
In conclusion, these studies implicate miR-21 in improving post-injurious outcomes in both preventative and therapeutic contexts. Overall, miR-21 shows great promise as a neuroprotective agent against stroke and possibly other acute CNS injuries.
Acknowledgements
This work was supported in part by the U.S. Department of Veterans Affairs Merit Review Grant I01-BX002985, and by National Institute of Health Grants RO1-144AAC1463 and RO1-144AAC1715. Mary Lopez was supported by a pre-doctoral Advanced Opportunity Fellowship through SciMed GRS at the University of Wisconsin-Madison.
References
Mary S. Lopez
1Cellular and Molecular Pathology Program, University of Wisconsin, Madison, WI, USA;
2Department of Neurological Surgery, University of Wisconsin, Madison, WI, USA
Robert J. Dempsey
1Cellular and Molecular Pathology Program, University of Wisconsin, Madison, WI, USA;
2Department of Neurological Surgery, University of Wisconsin, Madison, WI, USA
Raghu Vemuganti
1Cellular and Molecular Pathology Program, University of Wisconsin, Madison, WI, USA;
2Department of Neurological Surgery, University of Wisconsin, Madison, WI, USA;
3William S. Middleton Veteran’s Administration Hospital, Madison, WI, USA
Corresponding author
Raghu Vemuganti
Email:vemuganti@neurosurgery.wisc.edu
In a new window | Download PPT
Figure 1: miR-21 regulates apoptosis. Fas ligand (FasL), programmed cell death protein 4 (PDCD4), and phosphatase and tensin homolog (PTEN) are the best-characterized targets of miR-21. These proteins synergistically promote apoptosis. FasL is either membrane-bound or soluble and physically binds to the Fas death receptor (FasR) to initiate a caspase cleavage cascade resulting in apoptosis. PTEN is a tumor suppressor and a negative regulator of PI3K that phosphorylates PIP2 to form PIP3 that activates Akt pathway leading to cell death. As such, prevention of PTEN by miR-21 can lead to better cell survival following conditions like stroke and TBI. PI3K and Akt also modulate caspase-9 cleavage and hence, PTEN inhibition leads to prevention of apoptosis. Furthermore, PI3K/Akt activates mammalian target of rapamycin (mTOR) that inhibits PDCD4 activity. PDCD4 causes a protein expression bottleneck by inhibiting eukaryotic initiation factor 4a (eIF4a) and the transcription factor activator protein 1 (AP-1). Preventing eIF4a activity causes endoplasmic reticulum stress and inhibiting AP-1 prevents expression of beneficial genes such as vascular endothelial growth factor (VEGF). As inhibition of FasL, PTEN or PDCD4 together is a powerful strategy to prevent apoptosis and hence miR-21 is an attractive therapeutic to protect the post-injury brain. Tipped arrows indicate activation and blunted arrows indicate inhibition.
Metrics
| Full-Text | Supporting Information | ||
|---|---|---|---|
| Number | 12034 | 84 | 0 |
Copyright © 2017 Conditioning Medicine, All Rights Reserved.
Address: Conditioning Medicine Editorial Office, 3500 Terrace Street, Pittsburgh, PA, 15213, USA