Conditioning Medicine
International bi-monthly journal of cell signaling, tissue protection, and translational research.
Near infrared light amplifies endothelial progenitor cell accumulation after stroke
Time:2019-09-05
Number:12326
Author Affiliations

Conditioning Medicine 2019. 2(4):170-177.
Abstract
Damage-associated molecular pattern (DAMP) signals may play key roles in mediating non-cell autonomous effects of pre and post-conditioning. Here, we show that near-infrared (NIR) light stimulation of astrocytes increases calcium-dependent secretion of the prototypical DAMP, HMGB1, which may then accelerate endothelial progenitor cell (EPC) accumulation after stroke. Conditioned media from NIR-stimulated astrocytes increased EPC proliferation in vitro, and blockade of HMGB1 with siRNA diminished the effect. In vivo transcranial NIR treatment confirmed that approximately 40% of NIR could penetrate the scalp and skull. Concomitantly, NIR increased GFAP expression in normal mouse brain 30 min after the irradiation. In a mouse model of focal ischemia, repeated irradiation of NIR at days 5, 9, and 13 successfully increased HMGB1 in the peri-infarct cortex, leading to a higher accumulation of EPCs 14 days post-stroke. Conditioning and tolerance are now known to involve cell-cell signaling between all cell types in the neurovascular unit. Taken together, our proof-of-concept study suggests that NIR light may be an effective conditioning tool to stimulate astrocytic signaling and promote EPC accumulation after stroke.
Keywords: astrocytes, endothelial progenitor cells, near-infrared light, stroke.
Abstract
Damage-associated molecular pattern (DAMP) signals may play key roles in mediating non-cell autonomous effects of pre and post-conditioning. Here, we show that near-infrared (NIR) light stimulation of astrocytes increases calcium-dependent secretion of the prototypical DAMP, HMGB1, which may then accelerate endothelial progenitor cell (EPC) accumulation after stroke. Conditioned media from NIR-stimulated astrocytes increased EPC proliferation in vitro, and blockade of HMGB1 with siRNA diminished the effect. In vivo transcranial NIR treatment confirmed that approximately 40% of NIR could penetrate the scalp and skull. Concomitantly, NIR increased GFAP expression in normal mouse brain 30 min after the irradiation. In a mouse model of focal ischemia, repeated irradiation of NIR at days 5, 9, and 13 successfully increased HMGB1 in the peri-infarct cortex, leading to a higher accumulation of EPCs 14 days post-stroke. Conditioning and tolerance are now known to involve cell-cell signaling between all cell types in the neurovascular unit. Taken together, our proof-of-concept study suggests that NIR light may be an effective conditioning tool to stimulate astrocytic signaling and promote EPC accumulation after stroke.
Keywords: astrocytes, endothelial progenitor cells, near-infrared light, stroke.
Introduction
Original ideas of conditioning and tolerance were based on intra-cellular mechanisms whereby a sublethal stimulus triggers internal responses that protect the cell against subsequent damaging insults (Thuret et al., 2014; Xie et al., 2018). However, it is now more broadly recognized that beyond the single cell response, pre- and post-conditioning can modify complex cascades of cell-cell signaling between multiple cell types (Xing and Lo, 2017; Esposito et al., 2018; Jung et al., 2018). In the central nervous system (CNS), inflammatory crosstalk within the neurovascular unit could be critical for conditioning and tolerance (Adstamongkonkul and Hess, 2017; Islam et al., 2017; Yang et al., 2017; Bastian et al., 2018; Xie et al., 2018), and damage-associated molecular pattern (DAMP) mediators may play key roles (Gadani et al., 2015). For example, blockade of toll-like receptor increased neuronal survival after ischemia, whereas pre-conditioning with the receptor ligand including high-mobility group box 1 (HMGB1) decreased p38 MAPK signal-mediated injury after ischemia-reperfusion (Tang et al., 2007; Wang et al., 2016; Zhou et al., 2017). Therefore, finding ways to modulate DAMPs may be useful in the pursuit of conditioning therapies.
Within this context of conditioning crosstalk in the neurovascular unit, astrocytic responses may be especially relevant (Narayanan and Perez-Pinzon, 2017). Astrocytes comprise the most numerous non-neuronal cell type in the mammalian brain and play many biological roles, including regulation of blood flow, provision of nutrients to nervous tissue, and maintenance of extracellular ionic balance. After CNS injury, crosstalk between astrocytes and neuronal cells contribute to repair and recovery in the neurovascular unit (Iadecola and Nedergaard, 2007; Koehler et al., 2009; Belanger et al., 2011; Tsai et al., 2012). More recently, beyond cell-cell signaling within the brain per se, dynamic crosstalk between brain and systemic responses such as circulating blood cells may also be important in the pathophysiology of CNS injury or disease (Meisel et al. 2005; Offner et al. 2009). Our previous findings suggest that HMGB1 released from reactive astrocytes may promote endothelial progenitor cell (EPC) accumulation during stroke recovery (Hayakawa et al., 2012). Is it possible that conditioning therapies can be applied to amplify beneficial astrocyte signals?
Numerous preclinical studies have demonstrated that near-infrared (NIR) light may induce a broad range of biological effects including ATP generation, gene expression, and tissue healing (Fitzgerald et al., 2013; Yu et al., 2015; de Freitas and Hamblin 2016). Recently, it has been reported that NIR can be used to induce pre and post-conditioning (Agrawal et al., 2014; Liebert et al., 2017). To date, the majority of the proposed mechanisms involve intracellular mechanisms including modulation of mitochondria, regulation of anti-apoptotic and heat shock proteins, and promotion of anti-oxidant defense (Agrawal et al., 2014; Bartos et al., 2016; Liebert et al., 2017). Here, we seek to extend these ideas by asking whether NIR can also modulate inter-cellular DAMP signaling. In our proof-of-concept study, we assessed the ability of NIR to activate brain astrocytes and influence HMGB1-mediated EPC accumulation in the brain after stroke.
Materials and methods
Reagents: Ethylenediaminetetaacetic acid (EDTA), BAPTA-AM, and nicotinamide guanine dinucleotide sodium salt were purchased from Sigma-Aldrich.
Animals: One-day-old neonatal Sprague-Dawley (SD) rats and male C57bL/6 mice (24–27 g) were purchased from Charles River Laboratories (Wilmington, MA). All experiments were performed following an institutionally approved protocol in accordance with National Institutes of Health guidelines and the United States Public Health Service's Policy on Human Care and Use of Laboratory Animals and following Animals in Research: Reporting In vivo Experiments (ARRIVE) guidelines.(Kilkenny et al. 2010)
Near infrared (NIR) light: NIR light in an in vitro study was launched from Gallium Aluminum Arsenide (GaAlAs) diode with continuous wave having 808 nm wavelength (Marubeni, Tokyo, Japan). Before NIR irradiation, adequate calibration using the power detector was performed using a 24 well non-coated culture plate with a cover that could prevent light scattering to other wells. The cells were irradiated with a power density of 0, 10, 20, and 37 mW/cm2 for 2 mins to achieve energy densities of 0, 1.2, 2.4, and 4.4 J/cm2. In vivo transcranial NIR treatment was performed with the same device (Marubeni, Tokyo, Japan).
Primary astrocyte cultures: Primary astrocyte cultures were prepared from cerebral cortices of 2-day-old neonatal SD rats. Briefly, dissociated cortical cells were suspended in Dulbecco's modified Eagle medium (NBM, Life Technology) containing 25 mM glucose, 4 mM glutamine, 1 mM sodium pyruvate, and 10% fetal bovine serum, and plated on uncoated 25 cm2 flasks at a density of 6×105 cells/cm2. Monolayers of type 1 astrocytes were obtained 12-14 days after plating. Non-astrocytic cells such as microglia and neurons were detached from the flasks by shaking and removed by changing the medium. Astrocytes were dissociated by trypsinization and then reseeded on uncoated 6- and 24-well plates at a density of 1×105cells/cm2. After the cells reached 90-95% confluence (2-3 days after seeding), cultures were switched to serum-free EBM-2 medium containing 1% penicillin/streptomycin, and experiments were initiated 1 h later.
EPC isolation from rat spleen: Spleens were used for obtaining EPCs as previously described (Rosell et al., 2009; Hayakawa et al., 2012). For each independent experiment, spleens from 11-12 weeks old SD rats were kept in HBSS solution. Under the fume hood, spleens were mechanically minced, placed at 37°C for 15 min, and run through a 40-μm nylon membrane to obtain cell suspensions. Mononuclear cells (MNCs) were obtained by density gradient centrifugation with Ficoll-Paque Plus (Amersham Biosciences Corp, Buckinghamshire, UK). After a short wash with red blood cells lysis solution, isolated MNCs were gently washed twice with complete growth media EGM-2MV (Lonza, Basel, Switzerland). MNCs were finally resuspended in EGM-2MV and 3 × 107 MNCs per well were seeded on collagen I-coated six-well plates (Becton Dickinson Labware, San Jose, CA) and incubated in a 5% CO2 incubator at 37°C. The first media change was performed 3 days after plating. For each independent experiment early EPCs between days 5 and days 7 after seeding were used.
Conditioned media collection: To prepare astrocyte or EPC-conditioned media, we used cells at 90-95% confluence and grown in EBM-2 medium containing 1% penicillin/streptomycin for 24 hours. Astrocyte conditioned media (ACM), near infrared light-stimulated-astrocyte-conditioned media (NIR-ACM), EPC conditioned media (EPC CM), near infrared light-stimulated-EPC-conditioned media (NIR-EPC CM), or no cells conditioned media were collected 24 hours after laser stimulation. Conditioned media were collected and filtered using a 0.20-μm filter before use in EPC studies. All conditioned media were used without dilution in all experiments.
Cell proliferation Assays: Cell proliferation was assessed by WST reduction assay (Dojindo, Rockville, MD), which detects dehydrogenase activity of viable cells. The cells were incubated with 10% WST solution for 1 h at 37°C. Then the absorbance of the culture medium was measured with a microplate reader at a test wavelength of 450 nm and a reference wavelength of 630 nm.
Western blot analysis: To determine HMGB1 in astrocyte conditioned media, ACM or NIR-ACM was concentrated 30 times by using Vivaspin 500 centrifugal concentrators (Vivaproducts, Littleton, MA). Cultures were rinsed twice with ice-cold phosphate-buffered saline and the cells were collected into Pro-PREPTM Protein Extraction Solution (Boca Scientific, Westwood, MA). Each sample was loaded onto 4-20% Tris-glycine gels. After electrophoresis and transfer to nitrocellulose membranes (Novex), the membranes were blocked in Tris-buffered saline containing 0.1% Tween 20 and 0.2% I-block (Tropix) for 90 min at room temperature. Membranes were then incubated overnight at 4°C with the following primary antibodies, anti-GFAP antibody (1:1,000, BD Biosciences, San Jose, CA), anti-MAP2 antibody (1:1,000, Calbiochem, San Diego, CA), monoclonal anti-HMGB1 (1:2000, Abcam, Cambridge, MA), anti-CD38 antibody (1:500, Santa Cruz Biotechnology, Dallas, TX), anti-BDNF antibody (1:1000, Millipore, Burlington, MA), anti-IGF-1 antibody (1:700, Santa Cruz Biotechnology, Dallas, TX), anti-FGF-2 (1:700, Santa Cruz Biotechnology, Dallas, TX), anti-VEGF antibody (1:700, Santa Cruz Biotechnology, Dallas, TX). After incubation with peroxidase-conjugated secondary antibodies, visualization was enhanced by chemiluminescence (GE Healthcare, Chicago, IL). Optical density was assessed using NIH Image analysis software.
Fluorescence-activated cell sorting (FACS) analysis: FACS analysis was performed as described before (Hayakawa et al., 2012). Single cell suspensions were prepared by filtering through a 40-μm strainer. Cell suspensions were pre-blocked with 3% BSA and then incubated with the following primary antibodies against CD34 (1:100, GenWay Biotech, San Diego, CA), VEGFR2/Flk1/KDR (1:100, Abcam, Cambridge, MA). Fluorescent-tagged Fab specific secondary antibodies from Jackson laboratories were incubated for 30 min at room temperature. Labeled cell populations were measured by FACSCalibur (BD Biosciences, San Jose, CA). FACS data were analyzed by Cellquest pro software (BD Biosciences, San Jose, CA). FACS analysis were performed using a variety of controls including unstained samples, isotype antibodies, and single stained samples for determining appropriate gates, voltages, and compensations required in multivariate flow cytometry.
Immunocytochemistry and immunohistochemistry: Immunocytochemistry and immunohistochemistry were performed as described before (Hayakawa et al. 2010a; Hayakawa et al. 2010b). After staining with primary antibodies and fluorescent-tagged secondary antibodies, nuclei were counterstained with 4,6-diamidino-2-phenylindole (DAPI) and coverslipped. Immunostaining was analyzed with a fluorescence microscope (Olympus BX51, Waltham, MA) interfaced with a digital charge-coupled device camera and an image analysis system.
siRNA experiment in astrocyte culture: Control siRNA and HMGB1 siRNA were obtained from Santa Cruz Biotechnology, Inc. (Dallas, TX). Control siRNA (sc-37007) consisted of a scrambled sequence that would not lead to the specific degradation of any known cellular mRNA. HMGB1 siRNAs (sc-270015) were each a pool of 3 target-specific 19-25 nt siRNAs designed to knock down gene expression. The sequences for rat HMGB1 siRNAs were designed as followed: Sequence 1: 5’-GCAUAUUAGUACCAGUUGU-3’, Sequence 2: 5’-CUGCUUAGUUUAGGGAACA-3’, Sequence 3: 5’-GAGUCCUGGAUGAUACUAA-3’. siRNAs were prepared according to the transfection protocol for cell cultures from Santa Cruz Biotechnology. Briefly, 1 ml of siRNA transfection reagent mixture (Transfection reagent, sc-29528, Transfection medium, sc-36868) was co-incubated with astrocytes for 6 hours in a 5% CO2 incubator at 37°C and then the same amount of DMEM 20% FBS was added. An additional incubation was performed for 18 hours and then the procedure for conditioned media was carried on.
Mouse Focal Cerebral Ischemia Model: Male C57BL/6 mice (13-14 weeks) were anesthetized with 1% isoflurane and rectal temperatures and cerebral blood flow were monitored. After midline skin incision, a 7-0 nylon monofilament coated with silicon resin was introduced through a small incision into the common carotid artery. Adequate cerebral ischemia was assessed by Laser Doppler flowmetry and by examining forelimb flexion after the mice recovered from anesthesia. Forty-five mins after occlusion, the mice were re-anesthetized and reperfusion was established by withdrawal of the filament.
Statistical analysis: Results are expressed as mean ± SD. All experiments were performed with full blinding, allocation concealment, and randomization. When only two groups were compared, unpaired t-test was used. Multiple comparisons were evaluated by one-way ANOVA followed by Tukey test. P < 0.05 was considered to be statistically significant.
Results
Near-infrared (NIR) irradiation increased intracellular calcium and promoted HMGB1 secretion in rat cortical astrocytes.
As changes in intracellular calcium are critical to astrocyte function, we first used in vitro cell culture to measure these changes in response to NIR stimulation. Rat cortical astrocytes were irradiated with a power density of 0, 10, 20 and 37 mW/cm2 for 2 mins to achieve energy densities of 0, 1.2, 2.4, and 4.4 J/cm2. Fluo-4 AM-labeled intracellular Ca2+ increased in astrocytes after expose to NIR light (Figure 1a). Immunocytochemistry and western blot demonstrated that astrocytes secreted HMGB1 into the media (Figure 1b, c). We confirmed that blocking intracellular calcium signaling significantly suppressed HMGB1 secretion in NIR-stimulated astrocytes (Figure 1d-f).
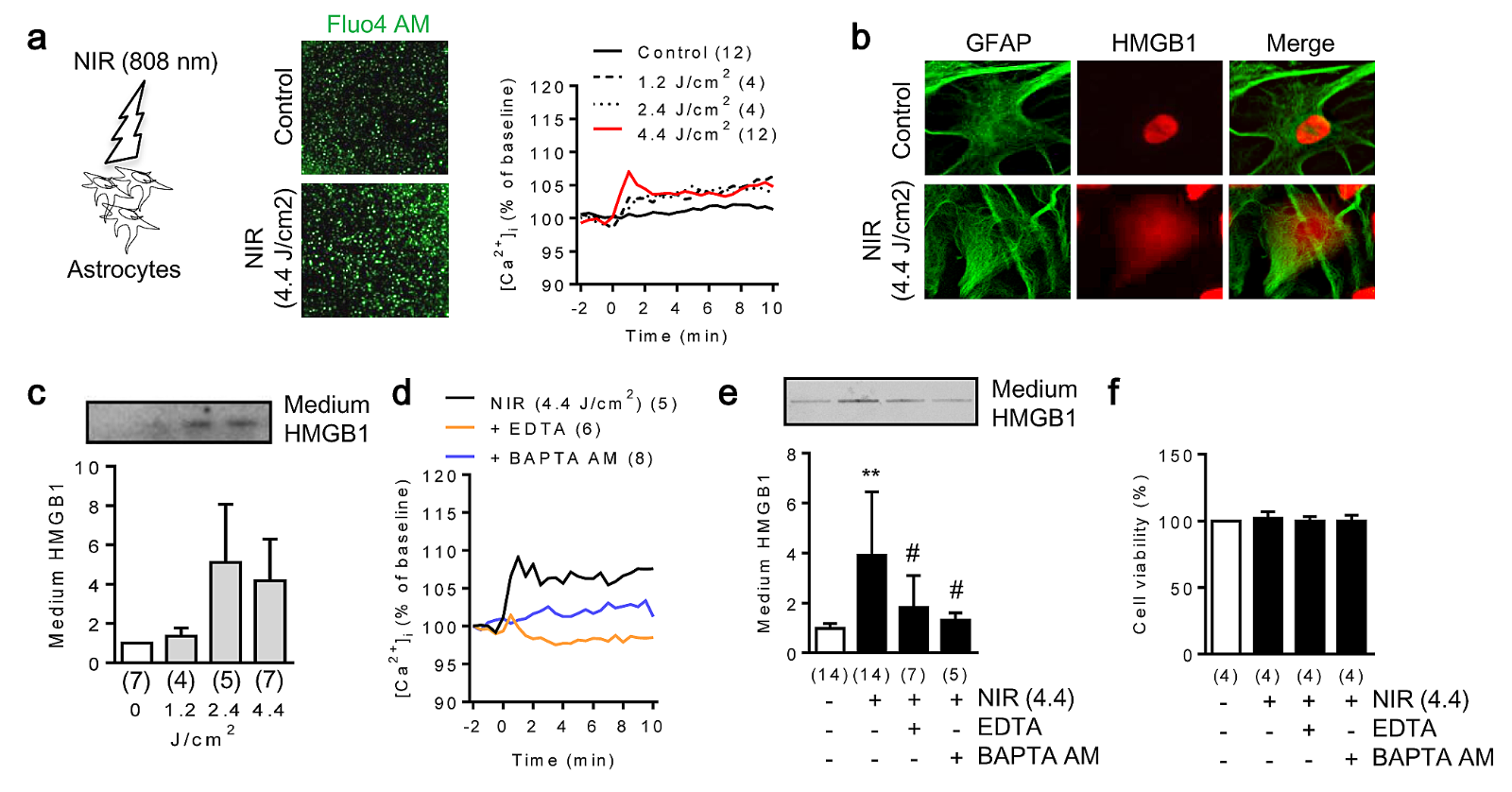
In a new window | Download PPT
Figure 1: Near infrared light (NIR) promotes HMGB1 secretion via an increase of intracellular calcium in rat cortical astrocytes. a. In vitro rat cortical astrocytes were exposed to NIR light (808 nm, 0, 10, 20, 37 mW/cm2 for 2 mins). Fluo-4 AM signal indicated intracellular calcium mobilization after NIR light therapy in astrocytes (n = 4-12). b. Immunocytochemistry demonstrated that HMGB1 was translocated from the nucleus to cell cytoplasm 1 hour after NIR irradiation. c. HMGB1 was released from astrocytes 24 hours after NIR irradiation (n = 4-7). d. Intracellular calcium mobilization after NIR stimulation was blocked by EDTA (10 μM) or BAPTA-AM (5 μM) (n = 5-8). e. Western blot analysis demonstrated that EDTA (10 μM) or BAPTA-AM (5 μM) significantly suppressed HMGB1 secretion mediated by NIR irradiation (n = 5-14). **p < 0.01 vs control, #p < 0.05 vs NIR (4.4). f. NIR did not influence cell viability (n = 4). All data are shown as mean ± SD.
NIR-stimulated astrocytes enhanced HMGB1-mediated EPC proliferation in vitro.
To test the idea that NIR-treated astrocytes may be a source of inter-cellular signals, we explored the effect of conditioned media from NIR-stimulated astrocytes (NIR-ACM) on EPC proliferation. Conditioned media from empty wells, control astrocytes (ACM), and stimulated astrocytes (NIR-ACM) were transferred to EPC cultures (Figure 2a). Interestingly, NIR-ACM remarkably enhanced EPC proliferation compared to ACM (Figure 2b, c). When endogenous HMGB1 in astrocytes was suppressed with siRNA (Figure 2d), the ability of NIR-ACM to enhance EPC proliferation was attenuated (Figure 2e). A caveat is whether NIR irradiation can directly increase HMGB1 and trigger autocrine responses in EPCs. To address this, NIR was used to irradiate EPCs, then conditioned media were collected from EPC cultures, and tested for EPC proliferation (Figure 2f). Conditioned media from empty wells, control EPCs (EPC CM), and stimulated EPCs (NIR-EPC CM) did not increase EPC proliferation, suggesting that NIR may not directly upregulate HMGB1 or induce proliferation in EPCs. Hence, the effects of NIR may be primarily mediated via astrocyte-EPC crosstalk.
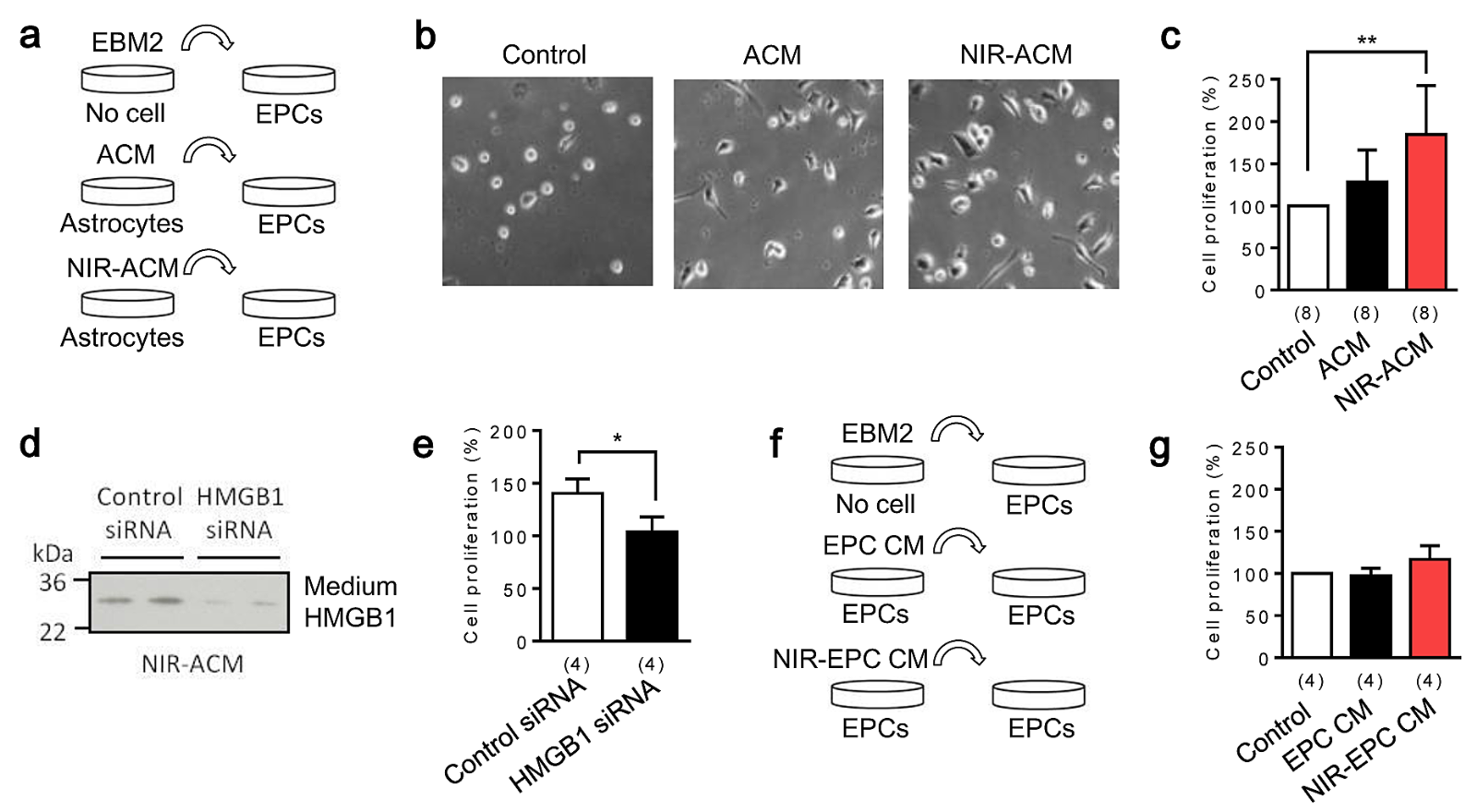
In a new window | Download PPT
Figure 2: Astrocytic HMGB1 increases endothelial progenitor cells (EPC) proliferation in vitro. a. Schematic experimental design for media transfer experiments. Conditioned media from an empty well (Control), normal astrocytes (ACM), and NIR-stimulated astrocytes (NIR-ACM) were collected 24 hours after incubation. Each conditioned media was transferred into early EPC cultures (day 5 after seeding) and incubated for 24 hours. b, c. NIR-ACM significantly enhanced early EPC proliferation compared with ACM (n = 8). **p < 0.01 vs control. d. A loss-of-function study was performed using siRNA. HMGB1 siRNA successfully suppressed HMGB1 secretion into conditioned media after NIR irradiation. e. NIR-ACM mediated EPC proliferation was significantly suppressed when astrocytic HMGB1 was decrease by siRNA (n = 4). *p < 0.05. f, g. Conditioned media collected from NIR-stimulated EPC did not increase EPC proliferation (n = 4). All data are shown as mean ± SD.
Transcranial NIR irradiation activates astrocytes in C57BL/6 mice.
Efficient NIR penetration through skin, skull, and the brain is well documented (Henderson and Morries 2015; Hart and Fitzgerald 2016). To confirm this in our own device and model systems, scalp and skull were dissected from mice to measure the percentage of NIR-light penetration (Figure 3a). NIR-light penetration was determined at three independent power irradiation levels. At all levels, we confirmed that ~40% of power was potentially detectable at the surface of cerebral cortex (Figure 3b, c). To test whether NIR irradiation activates astrocytes in vivo, we examined normal C57BL/6 male mice. Transcranial NIR treatment (808 nm, continuous wave, 37 mW/cm2 for 2 mins) was performed in a normal mouse after shaving the hair (Figure 3d), and then the brain was extracted 30 mins after treatment. Immunohistochemistry showed an enhancement of GFAP signal in the somatosensory cortex and the corpus callosum (Figure 3e). Western blot confirmed that GFAP protein levels in the cerebral cortex were significantly increased by transcranial NIR irradiation (Figure 3f). Taken together, these measurements support the penetration and biological feasibility of NIR in our in vivo mouse model.
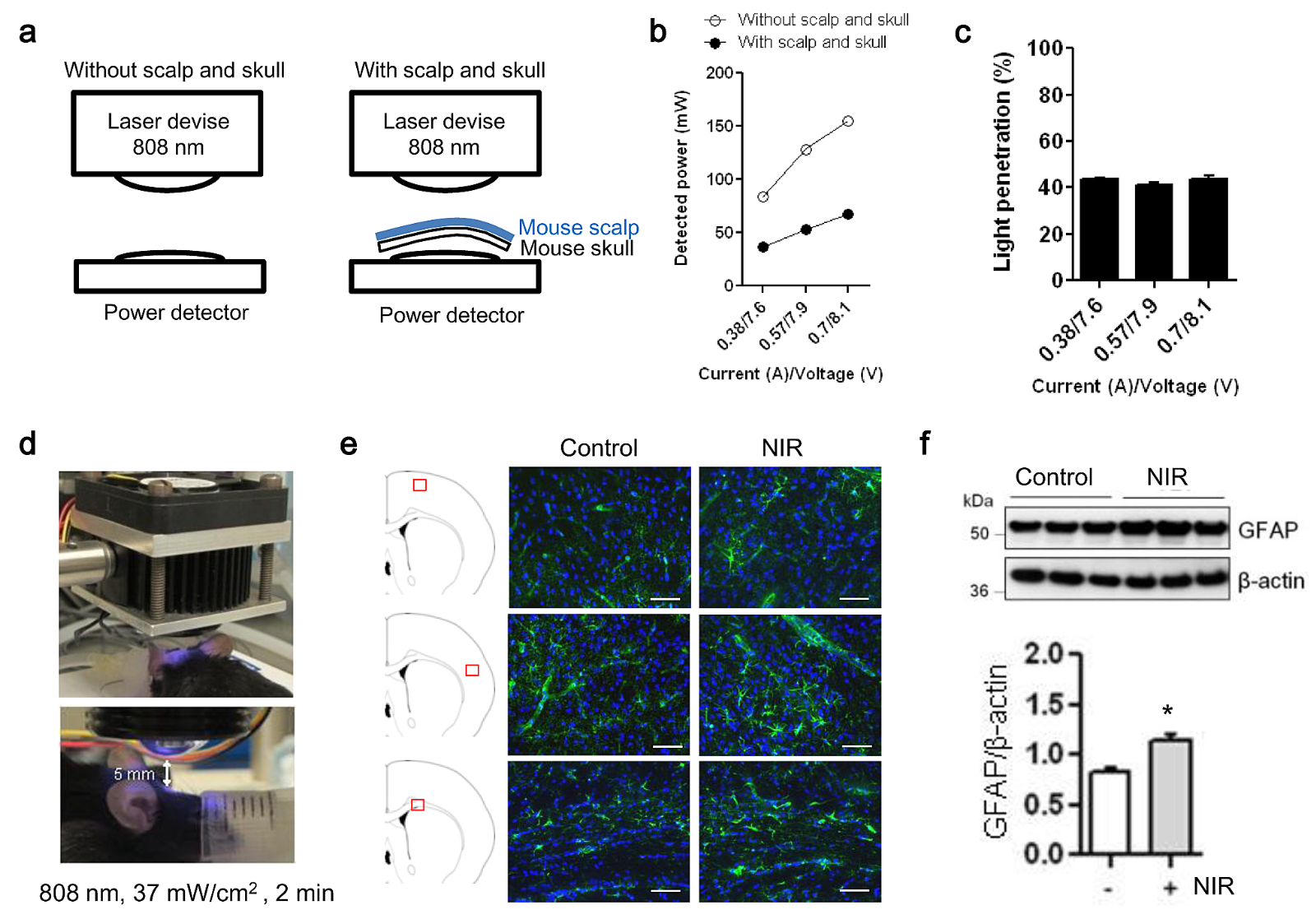
In a new window | Download PPT
Figure 3: In vivo transcranial NIR-light increases GFAP expression in the brains of treated C57BL/6 mice. a. Fresh scalp and skull were dissected from the mouse to measure a percentage of NIR-light penetration. b. NIR-light penetration was determined at three independent power irradiation levels. White circles show the power levels without mouse scalp and skull, and black circles show the powers reduced by fresh scalp and skull isolated from the mouse. c. With each irradiation, we confirmed that ~40% of power was reached into the surface of the cerebral cortex. d. Exposure to NIR light (808 nm, 37 mW/cm2 for 2 min) was performed with setting illuminator lens 5 mm from the mouse’s shaved scalp. e, f. GFAP positive astrocytes in primary somatosensory cortex and corpus callosum were determined by immunohistochemistry 30 min after NIR irradiation. Immunohistochemistry and western blot showed that compared to control, transcranial laser irradiation increased GFAP signal in both cerebral cortex and corpus callosum (n=3). Scale bar: 100 μm. *p<0.05 vs control. All data are shown as mean±SD.
Transcranial NIR irradiation increased HMGB1 expression and EPC accumulation in peri-infarct cortex after focal cerebral ischemia in C57bl6 mice.
To evaluate the effect of NIR irradiation in EPC accumulation in vivo, male C57BL/6 mice were subjected to transient focal cerebral ischemia and NIR was administered at 5, 9, and 13 days post-stroke (Figure 4a). After NIR irradiation, higher expression of HMGB1 was observed in reactive astrocytes in peri-infarct areas (Figure 4b). Western blot analysis confirmed up-regulation of HMGB1 in the peri-infarct cortex after NIR treatment (Figure 4c). Immunohistochemistry showed an accumulation of Flk1/CD34 double positive cells in somatosensory cortex in peri-infarct regions (Figure 4d). To further identify these cells, the percentage of Flk1/CD34 double-positive cells in cerebral blood vessel was measured by FACS. Flk1/CD34 double-positive cells in the lectin-positive endothelial population were operationally defined as EPCs in our model system as reported before (Hayakawa et al., 2012). With 3 rounds of NIR irradiation, EPCs were significantly increased in the peri-infarct cortex 14 days after stroke onset (Figure 4e).
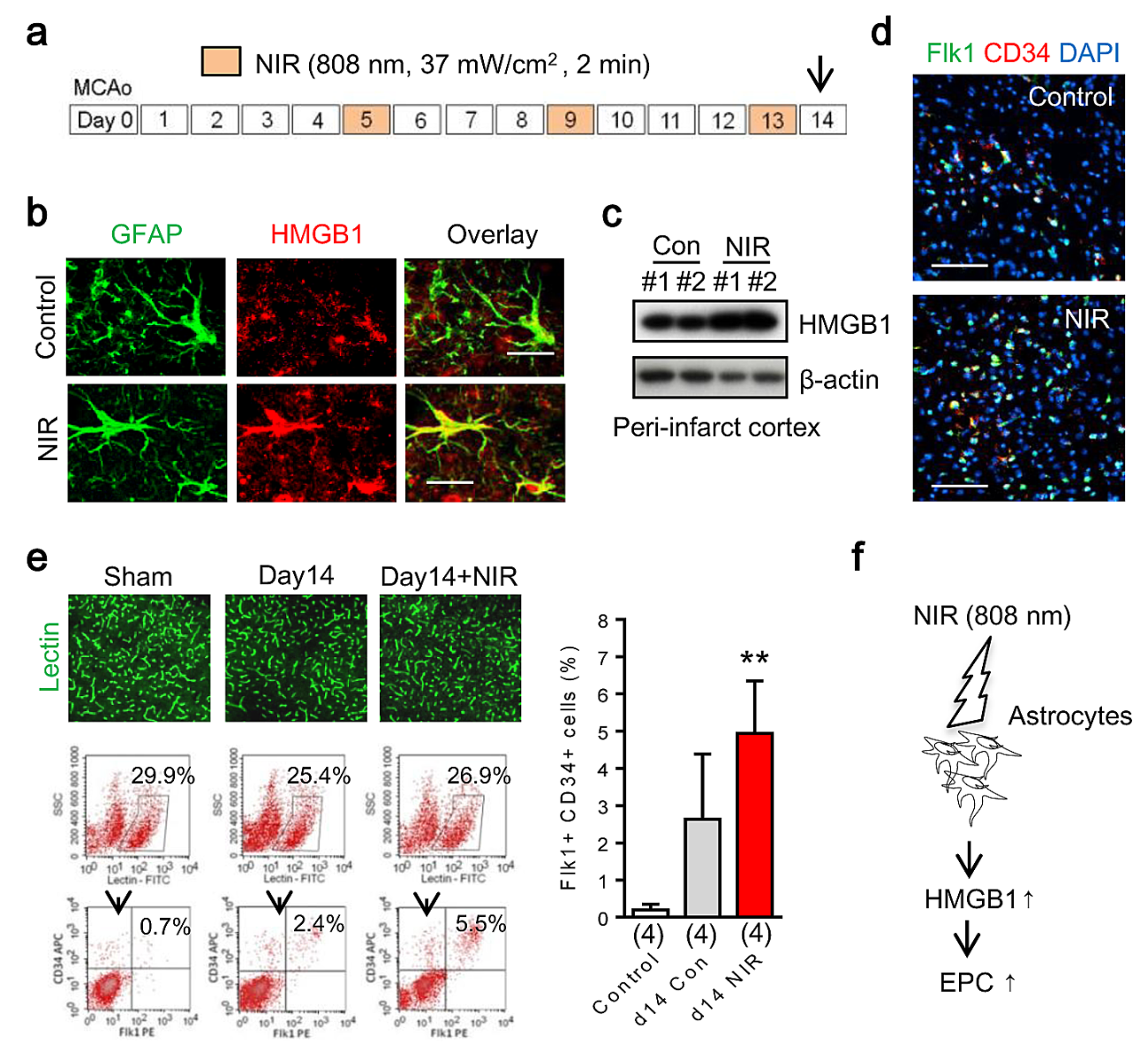
In a new window | Download PPT
Figure 4: In vivo transcranial near infrared light increases EPC accumulation after focal cerebral ischemia in C57bl6 mice. a. Exposure to NIR light (808 nm, 37 mW/cm2 for 2 min) was performed at days 5, 9, and 13 after stroke onset. b. NIR irradiation enhanced HMGB1 expression in GFAP positive reactive astrocyte in per-infarct area at day 14 after stroke. c. Western blot analysis confirmed that NIR upregulated HMGB1 in peri-infarct cortex. d. Immunohistochemistry showed an accumulation of Flk1/CD34 double-positive EPCs in somatosensory cortex in the peri-infarct region at day 14 after focal ischemia. e. FACS analysis demonstrated that with 3 rounds of NIR irradiation, EPCs were significantly increased in the somatosensory cortex 14 days after stroke onset. f. Transcranial NIR may become a tool to promote endogenous mechanisms of crosstalk between reactive astrocytes and EPCs after stroke.
Discussion
In summary, our in vitro and in vivo studies demonstrate that (i) astrocytes release HMGB1 after NIR light stimulation via intracellular calcium-dependent mechanisms, (ii) NIR-stimulated astrocytes promote HMGB1-mediated EPC proliferation in vitro, and (iii) transcranial NIR light treatment enhances endogenous astrocytic response and increases EPC accumulation after focal cerebral ischemia (Figure 4f). Collectively, these data suggest that NIR irradiation can be a conditioning tool to enhance crosstalk between astrocytes and EPCs after stroke and brain injury.
Cell–cell signaling between neuronal, glial, and vascular compartments provides the signals and substrates for investigating the mechanisms of acute injury and delayed recovery after stroke, brain injury and neurodegeneration (Iadecola 2004; Zlokovic 2010). Beyond cell-cell signaling within the brain per se, it has been proposed that dynamic crosstalk between the brain and circulating blood cells may also be important (Meisel et al. 2005; Offner et al. 2009). HMGB1, a highly conserved non-histone nuclear DNA-binding protein, is widely expressed in most eukaryotic cells (Yang et al., 2005), and can be passively released from damaged cells or actively secreted from stimulated cells in the brain (Kim et al., 2006; Hayakawa et al., 2008; Nakahara et al., 2009). Additionally, it has been reported that glial HMGB1 is crucial for EPC homing after ischemic insult (Hayakawa et al., 2012; Hayakawa et al., 2013; Chen et al., 2014; Hayakawa et al., 2014). What may be missing from the collective literature to date is how these endogenous responses can be boosted with non-invasive procedures. Our present study provides proof-of-principle that NIR irradiation may stimulate the endogenous astrocytic HMGB1 response and increase EPC accumulation after stroke.
However, there are a few caveats. Our data showed that NIR-stimulated astrocytes secreted HMGB1 via an intracellular calcium-dependent mechanism. But how the increase of intracellular calcium regulates HMGB1 secretion in astrocyte remains to be fully elucidated. Additional studies that dissect signaling pathways involved in HMGB1 secretion would be required to unequivocally prove causality. Second, NIR is known to be absorbed by mitochondria and influences intracellular bioenergetics and metabolism (Karu 1989; Morimoto et al., 1994). Dynamic changes in mitochondrial function after NIR irradiation may change overall astrocytic function including calcium homeostasis, ATP production, and the glutamate response (Jackson and Robinson 2018). Whether and how NIR influences astrocytic mitochondria besides HMGB1 secretion should be addressed in future study. Third, EPC-mediated outcomes following NIR treatment remain to be fully investigated. The collective literature demonstrates that EPC-derived soluble factors such as VEGF, HGF, and BDNF may induce acute neuroprotection, glial functional improvement, and promote oligodendrogenesis, neurogenesis, and angiogenesis (Liao et al., 2017; Maki et al., 2018; Santo et al., 2019). Our focus here is on the mechanistic interactions between NIR and EPCs. There is a large literature on the effects of NIR in animal models (Lapchak and Boitano 2016) but clinical trials have been negative (Hacke et al., 2014). Whether these NIR-EPC interactions can be leveraged to amplify beneficial outcomes should be addressed in future studies. Fourth, we proposed that HMGB1 was upregulated after NIR treatment after focal ischemia. However, HMGB1 is known as a biphasic DAMP molecule, i.e. HMGB1 may be deleterious in the acute stage of stroke (Qiu et al., 2010), while HMGB1 may also promote neurovascular remodeling via angiogenesis and neurogenesis in the delayed phases post-stroke (Huttunen et al., 2000; Hayakawa et al., 2012). Ultimately, a more rigorous NIR approach may be required in order to avoid endogenous deleterious responses mediated by HMGB1. Finally, our present study demonstrated that NIR enhanced the potential interaction between astrocytes and EPCs, but cell-cell interaction between NIR-stimulated astrocytes and other cell types remains to be fully explored. How NIR irradiation influences crosstalk signals in the brain need to be rigorously dissected.
Cell–cell signaling in the neurovascular unit provides the signals and substrates to restore brain function by the generation of new blood vessels, neurogenesis, and synaptogenesis in the late phase of injury (Iadecola 2004; Zlokovic 2010). Our findings demonstrate that beneficial effects of NIR within the neurovascular unit may be mediate in part via the promotion of astrocytic HMGB1 secretion and enhancement of EPC accumulation after stroke. Further studies are warranted to pursue and develop NIR as a non-invasive tool to modulate non-cell autonomous mechanisms of conditioning and tolerance in the CNS.
Acknowledgment:
This work was supported in part by grants from NIH and the Rappaport Foundation.
Competing Interests
The authors declare they have no competing financial interest.
References
Andrew Vahabzadeh-Hagh1,2
1Neuroprotection Research Laboratory, Departments of Radiology and Neurology, Massachusetts General Hospital and Harvard Medical School, Charlestown, Massachusetts, USA. 2Berenson-Allen Center for Noninvasive Brain Stimulation, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, Massachusetts, USA.
Thomas J. McCarthy3
3McCarthy Consultants, Inc., Boylston, Massachusetts, USA.
Luis De Taboada4
4LiteCure, LLC., New Castle, Delaware, USA.
Jackson Streeter5
5Quanterix Corporation, Inc., Billerica, Massachusetts, USA.
Alvaro Pascual-Leone2
2Berenson-Allen Center for Noninvasive Brain Stimulation, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, Massachusetts, USA.
Eng H. Lo1
1Neuroprotection Research Laboratory, Departments of Radiology and Neurology, Massachusetts General Hospital and Harvard Medical School, Charlestown, Massachusetts, USA.
Kazuhide Hayakawa1
1Neuroprotection Research Laboratory, Departments of Radiology and Neurology, Massachusetts General Hospital and Harvard Medical School, Charlestown, Massachusetts, USA.
Corresponding author:
Kazuhide Hayakawa
Email: Khayakawa1@mgh.harvard.edu
or
Eng H. Lo
Email: Lo@helix.mgh.harvard.edu
In a new window | Download PPT
Figure 1: Near infrared light (NIR) promotes HMGB1 secretion via an increase of intracellular calcium in rat cortical astrocytes. a. In vitro rat cortical astrocytes were exposed to NIR light (808 nm, 0, 10, 20, 37 mW/cm2 for 2 mins). Fluo-4 AM signal indicated intracellular calcium mobilization after NIR light therapy in astrocytes (n = 4-12). b. Immunocytochemistry demonstrated that HMGB1 was translocated from the nucleus to cell cytoplasm 1 hour after NIR irradiation. c. HMGB1 was released from astrocytes 24 hours after NIR irradiation (n = 4-7). d. Intracellular calcium mobilization after NIR stimulation was blocked by EDTA (10 μM) or BAPTA-AM (5 μM) (n = 5-8). e. Western blot analysis demonstrated that EDTA (10 μM) or BAPTA-AM (5 μM) significantly suppressed HMGB1 secretion mediated by NIR irradiation (n = 5-14). **p < 0.01 vs control, #p < 0.05 vs NIR (4.4). f. NIR did not influence cell viability (n = 4). All data are shown as mean ± SD.
In a new window | Download PPT
Figure 2: Astrocytic HMGB1 increases endothelial progenitor cells (EPC) proliferation in vitro. a. Schematic experimental design for media transfer experiments. Conditioned media from an empty well (Control), normal astrocytes (ACM), and NIR-stimulated astrocytes (NIR-ACM) were collected 24 hours after incubation. Each conditioned media was transferred into early EPC cultures (day 5 after seeding) and incubated for 24 hours. b, c. NIR-ACM significantly enhanced early EPC proliferation compared with ACM (n = 8). **p < 0.01 vs control. d. A loss-of-function study was performed using siRNA. HMGB1 siRNA successfully suppressed HMGB1 secretion into conditioned media after NIR irradiation. e. NIR-ACM mediated EPC proliferation was significantly suppressed when astrocytic HMGB1 was decrease by siRNA (n = 4). *p < 0.05. f, g. Conditioned media collected from NIR-stimulated EPC did not increase EPC proliferation (n = 4). All data are shown as mean ± SD.
In a new window | Download PPT
Figure 3: In vivo transcranial NIR-light increases GFAP expression in the brains of treated C57BL/6 mice. a. Fresh scalp and skull were dissected from the mouse to measure a percentage of NIR-light penetration. b. NIR-light penetration was determined at three independent power irradiation levels. White circles show the power levels without mouse scalp and skull, and black circles show the powers reduced by fresh scalp and skull isolated from the mouse. c. With each irradiation, we confirmed that ~40% of power was reached into the surface of the cerebral cortex. d. Exposure to NIR light (808 nm, 37 mW/cm2 for 2 min) was performed with setting illuminator lens 5 mm from the mouse’s shaved scalp. e, f. GFAP positive astrocytes in primary somatosensory cortex and corpus callosum were determined by immunohistochemistry 30 min after NIR irradiation. Immunohistochemistry and western blot showed that compared to control, transcranial laser irradiation increased GFAP signal in both cerebral cortex and corpus callosum (n=3). Scale bar: 100 μm. *p<0.05 vs control. All data are shown as mean±SD.
In a new window | Download PPT
Figure 4: In vivo transcranial near infrared light increases EPC accumulation after focal cerebral ischemia in C57bl6 mice. a. Exposure to NIR light (808 nm, 37 mW/cm2 for 2 min) was performed at days 5, 9, and 13 after stroke onset. b. NIR irradiation enhanced HMGB1 expression in GFAP positive reactive astrocyte in per-infarct area at day 14 after stroke. c. Western blot analysis confirmed that NIR upregulated HMGB1 in peri-infarct cortex. d. Immunohistochemistry showed an accumulation of Flk1/CD34 double-positive EPCs in somatosensory cortex in the peri-infarct region at day 14 after focal ischemia. e. FACS analysis demonstrated that with 3 rounds of NIR irradiation, EPCs were significantly increased in the somatosensory cortex 14 days after stroke onset. f. Transcranial NIR may become a tool to promote endogenous mechanisms of crosstalk between reactive astrocytes and EPCs after stroke.
Metrics
| Full-Text | Supporting Information | ||
|---|---|---|---|
| Number | 12326 | 19 | 0 |
Copyright © 2017 Conditioning Medicine, All Rights Reserved.
Address: Conditioning Medicine Editorial Office, 3500 Terrace Street, Pittsburgh, PA, 15213, USA