Browse Articles
Conditioning Medicine
International bi-monthly journal of cell signaling, tissue protection, and translational research.
Molecular mechanisms of intermittent fasting-induced ischemic tolerance
Time:2020-02-29
Number:15127
Author Affiliations

Conditioning Medicine 2020. 3(1): 9-17.
Abstract
Diet is a significant factor in determining human well-being. Excessive eating and/or diets with higher than needed amounts of carbohydrates, salt, and fat are known to cause metabolic disorders and functional changes in the body. To compensate the ill effects, many designer diets including the Mediterranean diet, the Okinawa diet, vegetarian/vegan diets, keto diet, anti-inflammatory diet, and the anti-oxidant diet have been introduced in the past 2 decades. While these diets are either enriched or devoid of one or more specific components, a better way to control diet is to limit the amount of food consumed. Caloric restriction (CR), which involves limiting the amount of food consumed rather than eliminating any specific type of food, as well as intermittent fasting (IF), which entails limiting the time during which food can be consumed on a given day, have gained popularity because of their positive effects on human health. While the molecular mechanisms of these 2 dietary regimens have not been fully deciphered, they are known to prolong the life span, control blood pressure, and blood glucose levels. Furthermore, CR and IF were both shown to decrease the incidence of heart attack and stroke, as well as their ill effects. In particular, IF is thought to promote metabolic switching by altering gene expression profiles leading to reduced inflammation and oxidative stress, while increasing plasticity and regeneration.
Keywords: Dietary restriction, Brain, Stroke, Neuroprotection, Inflammation, Oxidative stress.
Abstract
Diet is a significant factor in determining human well-being. Excessive eating and/or diets with higher than needed amounts of carbohydrates, salt, and fat are known to cause metabolic disorders and functional changes in the body. To compensate the ill effects, many designer diets including the Mediterranean diet, the Okinawa diet, vegetarian/vegan diets, keto diet, anti-inflammatory diet, and the anti-oxidant diet have been introduced in the past 2 decades. While these diets are either enriched or devoid of one or more specific components, a better way to control diet is to limit the amount of food consumed. Caloric restriction (CR), which involves limiting the amount of food consumed rather than eliminating any specific type of food, as well as intermittent fasting (IF), which entails limiting the time during which food can be consumed on a given day, have gained popularity because of their positive effects on human health. While the molecular mechanisms of these 2 dietary regimens have not been fully deciphered, they are known to prolong the life span, control blood pressure, and blood glucose levels. Furthermore, CR and IF were both shown to decrease the incidence of heart attack and stroke, as well as their ill effects. In particular, IF is thought to promote metabolic switching by altering gene expression profiles leading to reduced inflammation and oxidative stress, while increasing plasticity and regeneration.
Keywords: Dietary restriction, Brain, Stroke, Neuroprotection, Inflammation, Oxidative stress.
Introduction
Over the last century, caloric intake has steadily increased, but physical activity/exercise in humans has not. This has led to faster progression of metabolism-related disorders such as obesity, diabetes, and hypertension, and the associated diseases like heart attack, kidney dysfunction, and stroke. Recent studies have shown that caloric restriction (CR) extends lifespan, decreases age-induced functional changes, and minimizes the incidence of age-related diseases in multiple species (Wan et al., 2010; Ran et al., 2015; Ciobanu et al., 2017; Wahl et al., 2017). When subjected to focal ischemia, animals subjected to CR had smaller infarcts and better neurologic outcome (Varendi et al., 2014; Ran et al., 2015; Ciobanu et al., 2017). More importantly, CR induces ischemic tolerance in aged rodents as well (Manzanero et al., 2011). CR can be achieved by consuming <70% of suggested calories/day, which is challenging in the longer term. Intermittent fasting (IF) is a variation of CR in which a person can consume 100% calories, but only during an 8 to 10 h period/day. For example, a person can eat normally between 9 am to 6 pm and then go on a fast from 6 pm to 9 am the next day. This is much easier as ~8 of those 15 fasting hours are sleeping time. In addition, IF can be followed in a flexible format. That means, IF can be practiced on all days of a week with time-restricted fasting for 12 to 16 h or can be followed as alternate day fasting. IF was shown to produce similar effects to CR including extension of life span, attenuation of neurodegenerative and cardiovascular diseases, and increased cerebral plasticity (Fann et al., 2017). IF also decreases blood pressure and promotes better glucose handling in humans (Tikoo et al., 2007; van Bilsen et al., 2014; Mattson et al., 2017; Erdem et al., 2018; Sutton et al., 2018; Badreh et al., 2019 ; Camelo et al., 2019). In addition, conditioning by IF was shown to minimize neuronal death and improve neurological recovery if rodents were subjected to focal ischemia (Manzanero S et al., 2014; Arumugam et al., 2010). While minimizing the post-stroke brain damage with pharmacologic therapies is important; modalities that can prevent stroke incidence and/or promote tolerance (so that the brain damage is lower in the unavoidable event of a stroke) are also extremely beneficial. The goal of this review article is to discuss the putative mechanisms that confer ischemic tolerance following IF.
Decreasing food intake (caloric restriction and IF) is different than maintaining a designer diet. For example, to follow a designer diet like the Mediterranean diet or the anti-inflammatory diet, the person will eat a specific kind of food rich in certain ingredients, whereas, in CR/IF there are no specific foods to eat or avoid. A person can eat any food, but the amount consumed (CR) or time during which a person can eat (IF) is restricted. As of today, food restriction studies are not combined with any designer diets.
IF leads to extensive transcriptional reprogramming
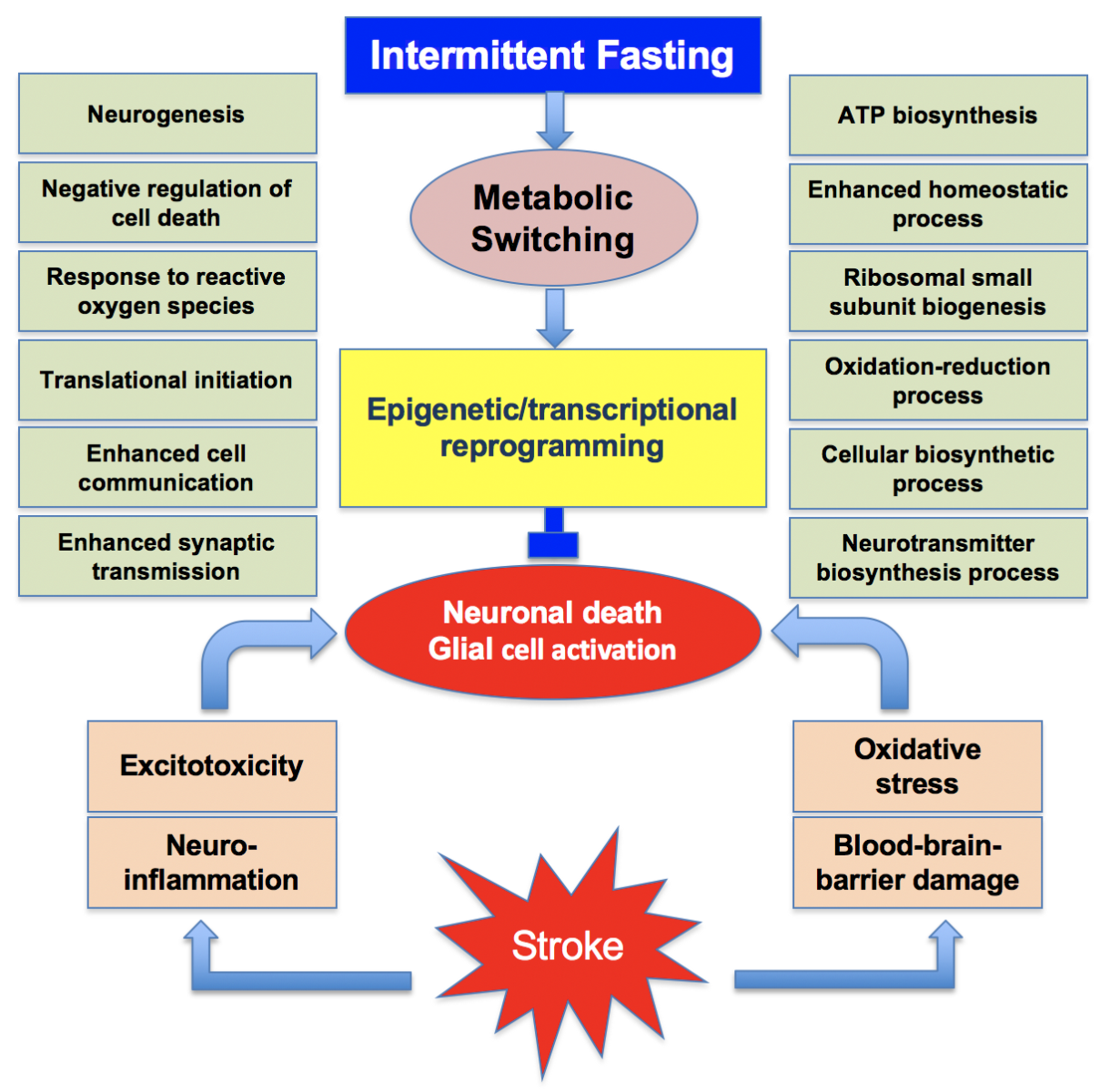
A recent study evaluated the detailed transcriptomic profiles in brain, heart, kidney, and muscle of mice subjected to various paradigms of IF (Kim et al., 2018). In this study, adult C57BL/6 male mice were subjected to either ad libitum (AL) feeding or IF for either 12h or 16h a day for 4 months. At the end of the IF period, the cerebral cortical tissue of the 3 cohorts of mice was analyzed by RNA-sequencing (RNA-seq). Interestingly, 16h IF, but not 12h IF, induced several changes in gene expression compared to AL control. Genes induced after IF include those that control signal transduction, cell communication, CNS development, positive regulation of metabolism, and cellular responses to stimuli. All these groups or genes are known to be beneficial. The authors also studied the effect of IF on stroke-induced gene expression changes. Focal cerebral ischemia was induced by transient middle cerebral artery occlusion (MCAO) in cohorts of mice subjected to either 12h or 16h IF or AL feeding. In these mice, the cerebral cortical transcriptome was assessed at 3h, 12h, 1 day, and 3 days of reperfusion. In the AL cohort of mice subjected to transient MCAO, genes that were involved in inflammation, cell stress response to cell death, apoptotic processes, and immune cell response/inflammation were observed to be significantly upregulated as expected. However, both 12h and 16h IF cohorts showed several post-MCAO changes compared to the AL cohort. In the IF groups subjected to focal ischemia, transcription, translation, oxidation-reduction process, response to reactive oxygen species, response to stress, ribosome biogenesis and assembly, cellular biosynthesis, oxidative phosphorylation, regulation of metabolism, and regulation of cell proliferation were the major biological processes upregulated compared to AL group. Furthermore, regulation of neuronal death, regulation of apoptosis, calcium signaling, glutamatergic signaling, cholinergic signaling, GABAergic signaling, dopaminergic signaling, MAPK signaling, mTOR signaling, and inflammatory mediator regulation were the major biological processes downregulated in IF/MCAO groups compared to AL/MCAO groups. In brief, IF led to molecular switching in the post-stroke brain that changed the balance towards a pro-cell survival and anti-inflammatory state (Fig. 1). Importantly, many genes that participate in pathways that mediate neuroplasticity, such as neurotrophic factors and neuronal energy metabolism such as peroxisome proliferator-activated receptors (PPARs) were upregulated in the IF groups compared to the AL groups. In addition, a metabolic shift from glucose to fatty acid oxidation and change in circadian rhythms was also seen in the IF groups (Kim et al., 2018).
In a new window | Download PPT
Figure 1: Many synergistic pathological events that include excitotoxicity, neuroinflammation, oxidative stress and blood-brain barrier damage promote the post-stroke brain damage. Intermittent fasting leads to metabolic switching that promotes epigenetic and transcriptional reprogramming. This induces the translation of many beneficial proteins concurrently inhibiting the translation of neurotoxic proteins. This promotes a neuroprotective phenotype.
Interestingly, other studies also found that IF improved glucose tolerance and altered the genes that control the circadian clock, aging, and autophagy in humans (Jamshed et al., 2019). When adult rats were subjected to IF for 1 week and then global cerebral ischemia was induced in them, they showed better recovery of cognitive function as assessed by a novel object recognition test and the Morris water maze test compared to the AL cohort 8 weeks after surgery (Hu et al., 2019). This study also found that the IF cohort of rats showed mitigated post-ischemic oxidative stress indicated by lower malondialdehyde levels and higher glutathione levels compared to the AL cohort. Furthermore, the IF group of rats showed higher expression levels of the anti-oxidant enzymes NADPH oxidase 1 and superoxide dismutase, and their upstream genes (Hu et al., 2019). This study further confirmed that IF leads to lower microglial activation and induced expression of sphingosine 1-phosphate receptor 1 leading to decreased post-ischemic inflammation compared with the AL group. In diet-induced obese mice, IF induced neuropeptide Y gene expression and increased norephinephrine levels in the hypothalamus leading to fat loss and retention of lean mass, which are beneficial to animals (Gotthardt et al., 2016). Serotonin controls feeding behavior in rodents and mammals, hence altering the serotonin transporter SERT can help to mitigate feeding related disorders. Severe IF (only 2 hours feeding per day) for 7 days in adult rats did not result in any changes in brain SERT expression, but did increase neuropeptide Y and proopiomelanocortin expression in the hypothalamus (Lauzurica et al., 2010). IF was also shown to induce anti-depressive effects by modulating the expression of certain genes. In mice subjected to 9 hours of fasting per day and administered imipramine, the p-CREB/CREB ratio in the hippocampus and frontal cortex was enhanced and the immobility time in the forced swim test was decreased (Li et al., 2014). The senescence-accelerated prone 8 (SAMP8) mice, which live shorter than normal mice, show many changes in the CNS, including decreased expression of sirtuin 1 and increased expression of forkhead box protein O1 (FoxO1) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB) targets leading to altered transcriptional control of genes related to cell proliferation and survival (Tajes et al., 2010). In addition, these mice also showed reduced levels of brain derived neurotrophic factor (BDNF) and heat shock protein 70 (HSP70). Interestingly 8 weeks of IF in SMAP8 mice showed recovery of sirtuin 1, BDNF, and HSP70 expression concomitant with decreases in acetyled NF-kB targets, c-Jun N-terminal kinase 1 (JNK1), and FoxO1 (Tajes et al., 2010). Interestingly, even a single episode of a 36h fast was shown to alter the expression of genes that control fatty acid oxidation, cell cycle, apoptosis, and inflammation in the blood of humans (Elliott et al., 2014). These studies showed that IF can promote better cell survival via altered gene expression leading to increased quality of life.
IF induced transcriptional reprogramming is not restricted to higher level organisms. IF has also shown to alter transcriptional regulation in Drosophila that shows youthful metabolic and behavioral switching in the CNS (Zhang et al., 2018). In C. elegans, IF led to altered expression of the gene Rheb-1, which is a low molecular weight GTPase that is responsible for controlling the insulin/insulin-like growth factor signaling effector DAP-16. Furthermore, in worms that were subjected to IF, RHEB-1 and target of rapamycin (TOR) signaling worked together to down-regulate insulin-like peptide INS-7. All these changes are thought to promote the life span in C-elegans subjected to IF (Honjoh et al., 2009).
IF protects the brain by curtailing inflammation
Post-stroke brain damage is known to be significantly modulated by central and peripheral inflammation that starts within hours after the insult (Dirnagl et al., 1999; Yi et al., 2007; Kapadia et al., 2008; del Zoppo, 2009; Iadecola and Anrather, 2011; Kim et al., 2014). A hallmark of inflammation after stroke is the extravasation of leukocytes (macrophages and neutrophils) from blood to brain parenchyma (Jin et al., 2010; Yang et al., 2019a). These cells release free radicals and pro-inflammatory molecules that mediate neuronal death in the ischemic brain. Previous studies showed that IF significantly mitigates several markers of inflammation in both the CNS and the periphery (Fann et al., 2017; Ott et al., 2017; Hu et al., 2019). Mechanistically, this was thought to be implemented by suppressing both genetic and physiological modulation.
In adult men, levels of the pro-inflammatory cytokines interleukin (IL)-6, IL1β, and tumor necrosis factor alpha (TNF-α), and the number of circulating leukocytes were observed to be lower during the Ramadan fasting period than 1 week before the start of the fast or 1 month after the fast (Faris et al., 2012). In a model of myocardial infarction, prior IF decreased leukocyte infiltration in the at risk area of the heart and the levels of the pro-inflammatory cytokine IL-6 in plasma (Wan et al., 2010). Lipopolysaccharide (LPS) administration is known to induce neuroinflammation in rodents that promotes cognitive deficits. This was observed to be due to NF-kB activation and thus induction of expression of many of its downstream pro-inflammatory genes including TLR4, iNOS, IL-1α, IL-1β, IFN-γ, TNF-α, and IL-6 (Vasconcelos et al., 2014). However, in rats subjected to IF for 30 days, LPS induced NF-kB activation, and downstream gene expression and cognitive deficits were observed to be curtailed compared to AL fed rats (Vasconcelos et al., 2014). This study lends further support to the idea that IF can rectify inflammatory gene expression following an insult in both the periphery and brain. Mechanistically, increased due to fasting leads to induction of sirtuins like SIRT1, which are deacetylases. SIRTs in turn activate the expression of downstream transcription factors including nuclear factor erythroid 2-related factor 2 (NRF2), FOXO, and PPARγ coactivator 1α (PGC-1α) (de Cabo and Mattson, 2019; Liu et al., 2019b; Mattson, 2019). These factors are known to induce anti-inflammatory and anti-oxidant gene expression. SIRT1 also prevents NF-kB activation by deacetylating the RelA/P65 subunit, thereby mitigating NF-kB downstream pro-inflammatory gene expression (Yeung et al., 2004). Another putative transcriptional mechanism that decreases inflammation is NAD+-mediated inactivation of mammalian TOR, which in turn promotes autophagy and mitophagy, thus decreasing pro-inflammatory cytokine levels (Fann et al., 2017).
Importantly, IF alleviated LPS-induced neuroinflammation in both young and aged rats, which was shown to be mediated by sodium-potassium adenosine triphosphatase (Na,K-ATPase) (Vasconcelos et al., 2015). IF, when combined with resistance training, significantly decreased the levels of pro-inflammatory cytokines IL-6, IL-1β, and TNF-α in the blood of adult male humans (Moro et al., 2016). Another study combined IF with supplementation of Ayurvedic herbs (leaf powder of Withania somnifera and dried stem powder of Tinospora cordifolia) in middle aged female rats. This combo treatment led to significantly reduced pro-inflammatory molecules IL-6, IL-1β, TNF-α, NF-kB, and Iba1 in the hippocampus (Singh et al., 2017). This study also observed that the rats subjected to IF + herbal supplements showed reduced anxiety-like behavior (Singh et al., 2017). These studies indicate that a combination of positive life style changes synergistically act to provide benefits to humans. In addition, IF was also protective in comorbid conditions. For example, when high-fat diet fed male mice were subjected to IF, there was a significant improvement in glucose tolerance and decreased fat mass associated with reduced levels of macrophage markers (Lgals3, Itgax, Ccl2, and Ccl3) indicating decreased inflammation (Liu et al., 2019a).
Inflammasomes are multimeric protein complexes, which are components of the innate immune system. When activated, they release several pro-inflammatory cytokines like IL-1β and IL-6 that promote cell death (Swanson et al., 2019). In particular, the NOD-, LRR- and pyrin domain-containing protein (NLRP) inflammasomes were shown to be activated by focal ischemia (Bauernfeind et al., 2009). Recent studies showed mice subjected to IF prior to stroke show reduced activation of NLRP inflammasomes, in addition to decreased levels of pro-inflammatory cytokines, which is beneficial for cells to survive after an ischemic attack (Fann et al., 2014). As inflammasomes are known to be activated by NF-kB, and as IF prevents NF-kB, it also leads to less inflammasome activation (Jung et al., 2009; Castello et al., 2010; Tajes et al., 2010; Fann et al., 2018). It was also shown that IF prevents post-ischemic activation of both NLRP1 and NLRP3 inflammasomes in mouse brain (Fann et al., 2014).
IF minimizes oxidative stress
Following stroke, white blood cells, including leukocytes and macrophages, that extravasate into brain parenchyma release several reactive oxygen species (ROS), free radicals (superoxide, hydroxyl radical, and peroxyl radical), and non-radicals (hydrogen peroxide and hyperchlorous acid). These cells also release reactive nitrogen species (RNS), radicals (nitric oxide and nitrogen dioxide), and non-radicals (peroxynitrite, dinitrogen trioxide, nitrous acid, nitroxyl anion, and nitrosyl cations), which are all neurotoxic (Sundararajan et al., 2005; Bowen et al., 2006; Kapadia et al., 2006; Satriotomo et al., 2006; Tureyen et al., 2007; Weston et al., 2007; Tang et al., 2008; Jin et al., 2010; Chu et al., 2014). Numerous studies have shown that IF before induction of stroke in rodent curtail release of ROS and RNS, as well as increase anti-oxidant defense by promoting the expression of enzymes like superoxide dismutase and catalase downstream to the transcription factor Nrf2 (Arumugam et al., 2010; Amigo and Kowaltowski, 2014 ; Ahn et al., 2018; Madkour et al., 2019). Fasting is known to cause metabolic switching resulting in lowered glucose and increased fatty acid use for energy production. IF promotes fatty acid production from triglycerides and the fatty acids are converted to ketone bodies that are used as a major source of fuel during IF (Mattson et al., 2017). Ketone bodies also modulate the expression of PGC-1α (Svensson et al., 2016). One of the mechanisms that is central to controlling oxidative stress after IF is thought to be initiated by the transcription factor PGC-1α, which promotes the levels of the antioxidant SIRT1 that in turn promotes the expression of mitochondrial uncoupling proteins (UCPs) UCP2 and UCP4 (Mattiasson et al., 2003; Bevilacqua et al., 2005; Liu et al., 2006; Canto and Auwerx, 2009; Chu et al., 2009; Haines et al., 2010; Canto and Auwerx, 2011). It was shown that when rodents were kept on IF prior to the induction of stroke, neuronal mitochondrial number and the levels of UCP2 and UCP4 were shown to be increased, which might lead to a higher rate of mitochondrial respiration (Fann et al., 2017). Overall, these effects decreased ROS levels, while concomitantly increasing their disposal leading to decreased oxidative stress after stroke.
IF is also thought to induce other transcriptional mechanisms that reduce oxidative stress. For example, IF activates the transcription factor NRF2, which induces many antioxidant enzymes including superoxide dismutase-2 and catalase. Furthermore, food restriction promotes mitochondrial biogenesis by activating transcription factors NRF2 and mitochondria transcription factor A (TFAM), and by modulating certain microRNAs (Hancock et al., 2011; Gouspillou and Hepple, 2013; Picca et al., 2013; Zhang et al., 2019). It was also suggested that IF might increase the availability of NAD+ due to mitochondrial uncoupling and also increase the respiratory rate that leads to reduced oxygen tension, subsequently lowering levels of superoxide production (Sanz et al., 2005; Haines et al., 2010; Fann et al., 2017; Klaus and Ost, 2019).
IF promotes brain plasticity
Rodents subjected to IF demonstrated improved cognitive function. Various studies with different regimens of IF showed improvement in verbal memory, spatial memory, working memory, associative memory, and executive function in adult as well as aged subjects (35 to 40 of Mattson). IF was shown to significantly increase the levels of BDNF, vascular endothelial growth factor (VEGF), and basic fibroblast growth factor (bFGF) in young as well as middle-aged male mice (Arumugam et al., 2010). BDNF is known to activate TrkB-PI3K/Akt signaling that induces neurogenesis in the adult brain following focal ischemia (Liu et al., 2015; Kisoh et al., 2017; Yang et al., 2019b). Hypoxia-induced factor 1 alpha (HIF-1α) induced by IF promotes the expression of VEGF-A, which might also increase neurogenesis. A recent study showed conclusive evidence for increased activation of the Notch signaling pathway, as well as upregulation of BDNF and downstream p-CREB following IF, which are all known to mediate neural stem cell proliferation (Baik et al., 2019). These authors further showed increased levels of PSD95, which is a post-synaptic marker, and nestin, which is an immature neuronal marker in the hippocampus of mice subjected to IF for 3 months (Baik et al., 2019). Hence, the improved cognitive function after IF might be due to neurotrophic factors that stimulate neurogenesis and angiogenesis. A diet that mimics fasting for 4 days resulted in an increased number of progenitors and stem cells leading to higher regeneration in adult mice (Brandhorst S et al., 2015). These authors also showed that a fasting mimicking diet promoted increased IGF-1 levels, hippocampal neurogenesis, and improved cognitive performance in old mice as well (Brandhorst S et al., 2015). It was also demonstrated that in amyloid precursor protein (APP) mutant mice IF enhanced hippocampal GABAergic tone leading to reduced anxiety-like behavior and improved hippocampus-dependent memory in a SIRT3 dependent manner (Liu et al., 2019b). Phosphatase and tensin homolog (PTEN) plays an essential role in modulating cell growth and survival. IF was shown to decrease impairment in contextual fear memory observed in PTEN haloinsufficient mice (Cabral-Costa JV 2018). A recent study showed that IF prevents TBI-induced hippocampal dependent learning and memory deficits in adult mice (Rubovitch et al., 2019).
Gap in our understanding of the clinical translation of IF for neuroprotection after stroke
Post-stroke brain damage is considered to be significantly dependent on age and sex. While at a younger age, males show higher brain damage and more severe loss of neurological function than females, this trend reverses at older age. Post-menopausal women show higher incidence of stroke and importantly more brain damage and significantly worsened neurological dysfunction than men at older ages (Li and McCullough, 2009; Liu et al., 2009b; Liu et al., 2009a; McCullough et al., 2009; Liu and McCullough, 2012). Hence, the efficacy of IF in protecting the brain after stroke as well as other conditions like TBI, spinal cord injury, and chronic neurodegenerative disorders need preclinical testing in both sexes and more importantly using older animals.
As the consumption of processed foods and calorically-rich foods with higher amounts of fats and sugars increased steadily in humans and the level of physical activity decreased, the incidence of metabolic syndromes like type-2 diabetes, obesity, and hypertension increased at an alarming rate. These changes are known to exacerbate the risk of CNS and peripheral organ diseases enormously. In particular, type-2 diabetes and hypertension are the major comorbid conditions that promote the incidence of stroke as well as exacerbated post-stroke brain damage. Hence, future studies need to test the efficacy of IF in comorbid animal models like diabetic and hypertensive rodents.
Although many diabetic and hypertensive individuals are on medications, diet plays a major role in deciding the health of these patients. Adopting either CR or IF was shown to promote better glycemic homeostasis in diabetic individuals (Mattson et al., 2017; Anton et al., 2018 ; Zubrzycki et al., 2018). In humans, IF was shown to decrease body weight, fasting glucose, and insulin sensitivity in overweight adults (Barnosky et al., 2014). IF was shown to result in better glycemic control in type-2 diabetic humans (Carter et al., 2016a,b). More importantly, in type-2 diabetes patients, IF was shown to be a safe and tolerable practice that promotes better fasting glucose levels (Arnason et al., 2017). In prediabetic humans, IF was shown to improve insulin sensitivity and β cell responsiveness, while decreasing oxidative stress and blood pressure (Sutton et al., 2018). A recent study showed that intermittent CR in db/db mice resulted in significantly reduced fasting blood glucose levels and improved insulin sensitivity (Wei et al., 2019). IF was also shown to prevent progression of nephropathy in type 1 diabetic rats (Tikoo et al., 2007) and prevented hyperglycemia in diabetes-susceptible New Zealand Obese mice (Baumeier et al., 2015). Mechanistically, IF was shown to enhance beta cell survival in diabetic mice by preserving organelle quality via the autophagy-lysosome pathway (Liu et al., 2017). IF was also shown to promote neogenesis of beta cells by autophagy-induced degradation of Notch1 leading to reversal of type-2 diabetes symptoms in rodents (DiNicolantonio and McCarty, 2019).
IF was also shown to decrease blood pressure in young and aged rodents (Badreh et al., 2019 ), as well as rodents on high-fat or high-fructose diets (Camelo et al., 2019). More importantly, IF was also shown to reduce blood pressure in humans with hypertension (Erdem et al., 2018) and or pre-diabetes (Sutton et al., 2018).
The mechanisms by which IF promotes ischemic tolerance is not yet completely deciphered. Recent studies showed that the gut microbiome influences the physiology of many organs including the brain. Stroke was shown to significantly alter the composition of the gut microbiome that significantly influences the post-stroke outcome. Furthermore, the gut microbiome depends on eating habits, and hence IF influences the composition of microbes in the gut. Hence, future studies are needed to understand the effect of IF on the gut microbiome in both sexes at different ages and also in comorbid subjects. The gut and the brain communicate with each other and the microbiota is a major mediator of the “gut-brain axis”. An imbalance in this axis is thought to mediate several disorders including brain dysfunction. Dysbiosis of the gut microbiome is currently thought to be a major proponent of inflammation and brain damage after stroke (Benakis et al., 2016; Singh et al., 2016; Spychala et al., 2018; Durgan et al., 2019; Park et al., 2019; Prame Kumar and Wong, 2019). The gut microbiota is known to release several short chain fatty acids (SCFAs) that play key roles in protecting the brain after stroke (Zeng and Cullen, 2004; Chen et al., 2019). A recent study showed that supplementation of SCFAs altered microglial activation and improved post-stroke motor function recovery in mice subjected to focal ischemia (Sadler et al., 2019). Several studies showed that CR impacts the gut microbial composition and impacts metabolism (Zhang et al., 2013; Duszka et al., 2018; Fabbiano et al., 2018; Zheng et al., 2018). CR was shown to promote reduction of the firmicutes/bacteriodetes ratio and expansion of lactobacilli leading to increased short-chain fatty acid synthesis, which is beneficial (Tanca et al., 2018). CR-mediated gut microbiome changes were shown to promote anti-inflammatory effects in rodents (Pan et al., 2018), as well as humans (Ott et al., 2017). These actions were thought to be due to expansion of lactobacillus in the gut (Fraumene et al., 2018). IF was also shown to decrease obesity and insulin resistance probably by shifting gut microbiota composition to a beneficial phenotype (Li et al., 2017). In type-2 diabetic db/db mice, IF increased lifespan, in addition to decreasing acellular capillaries and leukocyte infiltration (Beli et al., 2018). Furthermore, IF in db/db mice altered the gut microbiome with increased levels of firmicutes and decreased levels of bacteroidetes and verrucomicrobia, which is thought to be a beneficial phenotype (Beli et al., 2018). This beneficial phenotype of gut microbiota was also shown to be associated with increased levels of tauroursodeoxycholate (TUDCA), which is a neuroprotective bile acid formed by firmicutes (Beli et al., 2018). IF protects the brain from autoimmune conditions by altering the gut microbiota (Cignarella et al., 2018). Composition of gut microbiota also correlates with immune function (Li et al., 2019). Hence, future studies are needed to decipher if the beneficial effects of IF after stroke are due to rectifying the gut microbiota dysbiosis.
Conclusions
Overall, we conclude that IF promotes an adoptive (stress) response in the body that includes prevention of inflammation, better handling of oxidative stress, formation of more mitochondria, transcriptional switch to turn off neurotoxic and to turn on neuroprotective genes, as well as increase brain plasticity through neurogenesis/angiogenesis. IF was shown to protect mature neurons, and to promote regeneration and plasticity by inducing neurogenesis. Most of the studies to date indicated a beneficial effect of IF in various species. An attractive feature for adoption of IF in humans is the flexibility, as the regimen can be followed for 14 to 16 hours/day or fasting on alternate days or even normal feeding on 5 days and reduced calories on 2 days of the week. All of these adaptations are known to be beneficial by reducing the incidence and negative effects of major metabolic disorders like diabetes and hypertension, as well as subsequent diseases like heart attack, stroke, or neurodegenerative diseases.
References
Raghu Vemuganti1,2
1Department of Neurological Surgery, University of Wisconsin, Madison, WI, USA. 2William S. Middleton VA Hospital, Madison, WI, USA.
Thiruma V. Arumugam3
3Department of Physiology, National University of Singapore, Singapore.
Corresponding author:
Raghu Vemuganti
Email: vemuganti@neurosurgery.wisc.edu
In a new window | Download PPT
Figure 1: Many synergistic pathological events that include excitotoxicity, neuroinflammation, oxidative stress and blood-brain barrier damage promote the post-stroke brain damage. Intermittent fasting leads to metabolic switching that promotes epigenetic and transcriptional reprogramming. This induces the translation of many beneficial proteins concurrently inhibiting the translation of neurotoxic proteins. This promotes a neuroprotective phenotype.
Metrics
| Full-Text | Supporting Information | ||
|---|---|---|---|
| Number | 15127 | 81 | 0 |
Copyright © 2017 Conditioning Medicine, All Rights Reserved.
Address: Conditioning Medicine Editorial Office, 3500 Terrace Street, Pittsburgh, PA, 15213, USA